

خلاصه
حفاظت از تنوع زیستی زمینی ; تحلیل کمیت عود (RQA) ; تبخیر و تعرق (ET)
1. معرفی
2. مواد و روشها
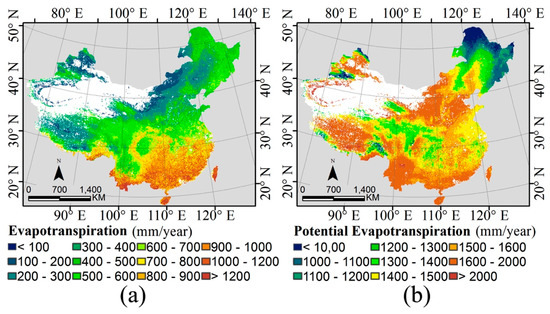
2.1. دینامیک آب-انرژی و مجموعه داده های سنجش از دور
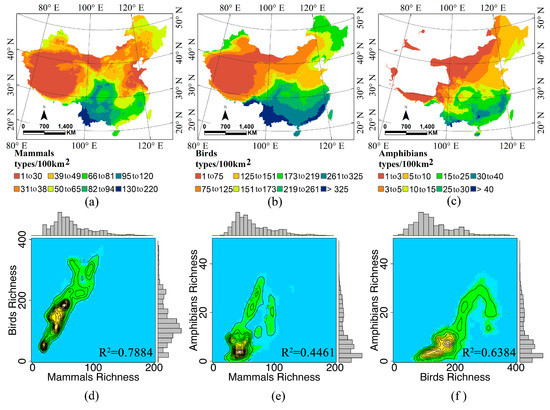
2.2. غنای گونه ها و مجموعه داده های سنجش از دور
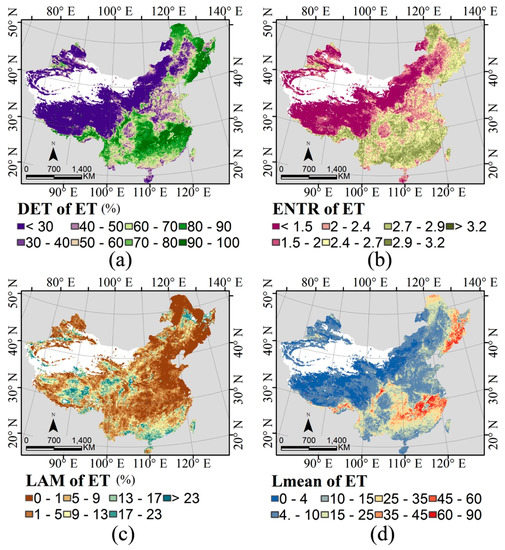
2.3. شاخص های پایداری زمانی
PET و ET منعکس کننده پویایی آب و هوا و نحوه تعامل آن با اکوسیستم هستند. با این حال، دینامیک آب و هوا و اکوسیستم ذاتاً در مکان و زمان غیر خطی هستند [ 13 ، 24 ]. بنابراین، مهم است که در نظر بگیریم که چگونه ثبات زمانی مجموعه دادههای سری زمانی بر گونهها تأثیر میگذارد. در این مطالعه، جبرگرایی (DET)، آنتروپی طول مورب (ENTR)، طول قطری متوسط (Lmean) و لایهپذیری (LAM) با استفاده از تحلیل کمیت عود (RQA، برای جزئیات بیشتر به [25] مراجعه کنید) برآورد شد . این شاخصها قبلاً برای توصیف دادههای سری زمانی بهدستآمده از سیستمهای اجتماعی به سیستمهای بیولوژیکی استفاده شدهاند [ 13 ، 24]]. در اینجا، از این چهار شاخص برای ارزیابی پایداری زمانی سری ET استفاده شد. این چهار شاخص بر اساس ساختار خط مورب و عمودی نمودار عود (RP، که به عنوان یک مسیر رسم می شود) است. ایکسمن→∈آرn ( i = 1 ، … ، N)ایکسمن→∈آر� (من=1،…،ن)در فضای فاز n بعدی و نشان دهنده بازگشت مسیر فضای فاز به یک حالت خاص از طریق یک نمودار ماتریسی است، که در آن سفید = 0 و سیاه = 1. دو محور در RP نشان دهنده زمان است [25، 26 ] ) . در هیستوگرام، خطوط مورب (نماینده تکامل محلی مشابه قسمتهای مختلف مسیر) طولی برابر با l دارند، در حالی که خطوط عمودی (که نشاندهنده یک حالت بدون تغییر برای مدتی است) دارای طول v در RP هستند، با تعداد کل زیر آن در [ 25 ]:
جایی که ε�آستانه فاصله است.
DET جبر (پیشبینیپذیری) یک سیستم معین را اندازهگیری میکند و نسبت نقاط بازگشتی است که ساختارهای مورب را به همه نقاط بازگشتی تشکیل میدهند. DET از 0 تا 1 است که در آن مقادیر نزدیک به 0 نشان دهنده یک سری تصادفی است، در حالی که مقادیر نزدیک به 1 نشان دهنده یک سری قطعی [ 25 ] است.
ENTR به آنتروپی شانون احتمال ( p(l) ) برای یافتن یک خط مورب با طول دقیق l در RP اشاره دارد و پیچیدگی RP را در رابطه با خطوط مورب منعکس می کند. ENTR بالا نشان دهنده پیچیدگی بالا است [ 25 ].
LAM که لامیناریته نامیده می شود، نسبت بین عود نقاطی است که ساختارهای عمودی را تشکیل می دهند و کل مجموعه نقاط عود. LAM مقدار حالتهای آرام را اندازهگیری میکند (انتقال دورهای-آشوب/آشوب-پریودیک، و همچنین انتقال آشوب-آشوب) [ 25 ، 26 ].
Lmean میانگین طول خط مورب است، که میانگین زمانی است که دو بخش از مسیر به یکدیگر نزدیک هستند و میانگین زمان پیشبینی را نشان میدهد [ 25 ].
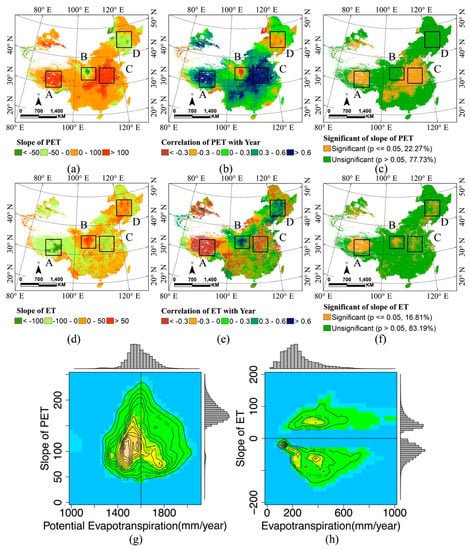
2.4. تحلیل گرایش
روندها با داده های شبکه ای در مورد پیش بینی کننده های احتمالی غنای گونه ها برای کشف علل بالقوه تغییرات مشاهده شده تجزیه و تحلیل شدند. برای تخمین دینامیک PET و ET بلند مدت (2000-2013)، یک تحلیل رگرسیون خطی ساده با حداقل مربعات [ 27 ] برای PET و ET سالانه اعمال شد.
که در آن t ∈ {2000، 2001،…، 2013} سال خاص را نشان می دهد، T تعداد کل سال ها، و DN تعداد دیجیتال سلول های شبکه PET یا ET برای یک سال خاص است. روند PET یا ET زمانی که شیب مثبت باشد افزایش می یابد و بالعکس. این تجزیه و تحلیل با استفاده از تابع ‘REGRESS’ در IDL©6.4 (زبان داده های تعاملی) و ENVI©4.4 (محیط برای تجسم تصاویر) انجام شد.
2.5. تحلیل آماری
3. نتایج و بحث
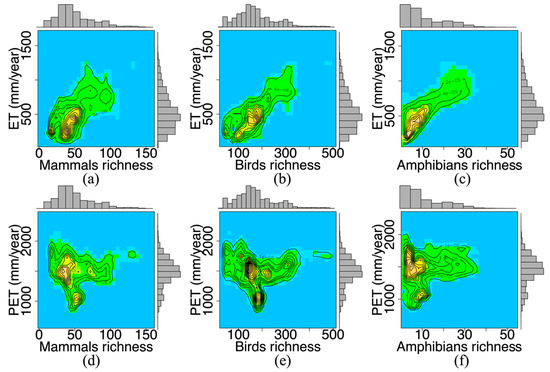
3.1. دینامیک آب-انرژی و غنای گونه ها
3.2. ثبات زمانی و غنای گونه ای
3.3. گرایش و تغییر
4. نتیجه گیری
منابع
- Mantyka-pringle، CS; مارتین، تی جی; رودز، JR تعاملات بین اقلیم و اثرات از دست دادن زیستگاه بر تنوع زیستی: یک بررسی سیستماتیک و متاآنالیز. گلوب. چانگ. Biol. 2012 ، 18 ، 1239-1252. [ Google Scholar ] [ CrossRef ]
- کیس، ام جی. لاولر، جی جی. توماسویچ، JA حساسیت نسبی به تغییرات آب و هوایی گونه ها در شمال غربی آمریکای شمالی. Biol. حفظ کنید. 2015 ، 187 ، 127-133. [ Google Scholar ] [ CrossRef ]
- فریتز، SA; Bininda-Emonds، OR; Purvis, A. تنوع جغرافیایی در پیش بینی خطر انقراض پستانداران: بزرگ بد است، اما فقط در مناطق استوایی. Ecol. Lett. 2009 ، 12 ، 538-549. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- هاوکینز، کارشناسی; فیلد، ر. کرنل، HV; کری، دی جی; Guégan، J.-F. کافمن، دی.م. کر، جی تی؛ Mittelbach، GG; اوبردورف، تی. O’Brien، EM انرژی، آب، و الگوهای جغرافیایی در مقیاس وسیع غنای گونه ها. اکولوژی 2003 ، 84 ، 3105-3117. [ Google Scholar ] [ CrossRef ]
- ژانگ، ک. کیمبال، جی اس. مو، کیو. جونز، لس آنجلس; گوتز، اس جی. در حال اجرا، تجزیه و تحلیل مبتنی بر ماهواره SW از روند شمال ET و تغییرات مرتبط در تعادل آب منطقه ای از سال 1983 تا 2005. J. Hydrol. 2009 ، 379 ، 92-110. [ Google Scholar ] [ CrossRef ]
- O’Brien، EM نسبیت بیولوژیکی به دینامیک آب-انرژی. J. Biogeogr. 2006 ، 33 ، 1868-1888. [ Google Scholar ] [ CrossRef ]
- O’Brien، E. دینامیک انرژی آب، آب و هوا، و پیش بینی غنای گونه های گیاهی چوبی: یک مدل کلی موقت. J. Biogeogr. 1998 ، 25 ، 379-398. [ Google Scholar ] [ CrossRef ]
- گیلمن، LN; رایت، SD؛ کوزنز، جی. مک براید، پی.دی. ملحی، ی. Whittaker، RJ Latitude، بهره وری و غنای گونه ای. گلوب. Ecol. Biogeogr. 2015 ، 24 ، 107-117. [ Google Scholar ] [ CrossRef ]
- هاوکینز، کارشناسی; پورتر، EE تأثیرات نسبی عوامل فعلی و تاریخی بر الگوهای تنوع پستانداران و پرندگان در آمریکای شمالی یخ زدایی شده. گلوب. Ecol. Biogeogr. 2003 ، 12 ، 475-481. [ Google Scholar ] [ CrossRef ]
- رودریگز، MA; Belmontes، JA انرژی، آب و الگوهای در مقیاس بزرگ از غنای گونه های خزندگان و دوزیستان در اروپا. Acta Oecol. 2005 ، 28 ، 65-70. [ Google Scholar ] [ CrossRef ]
- Whittaker، RJ; نوگوس-براوو، دی. Araújo، MB شیب جغرافیایی غنای گونه ای: آزمون حدس انرژی آب هاوکینز و همکاران. (2003) با استفاده از داده های اروپایی برای پنج گونه. گلوب. Ecol. Biogeogr. 2007 ، 16 ، 76-89. [ Google Scholar ] [ CrossRef ]
- کری، دی جی; Mittelbach، GG; کرنل، HV; فیلد، ر. Guégan، JF; هاوکینز، کارشناسی; کافمن، دی.م. کر، جی تی؛ اوبردورف، تی. O’Brien، E. پیشبینیها و آزمونهای فرضیههای مبتنی بر آب و هوا در مورد تنوع در مقیاس وسیع در غنای طبقهبندی. Ecol. Lett. 2004 ، 7 ، 1121-1134. [ Google Scholar ] [ CrossRef ]
- پرولکس، آر. پاروت، ال. فهریگ، ال. کری، دی جی تکرارهای طولانی مدت در اکولوژی: تشخیص روابط بین پویایی آب و هوا و تنوع زیستی در امتداد یک گرادیان عرضی. در تحلیل کمیت تکراری ; Springer: برلین، آلمان، 2015; صص 335-347. [ Google Scholar ]
- بینی، LM; دینیز فیلهو، JAF; هاوکینز، BA توضیحات ماکرواکولوژیکی برای تفاوت در شیب غنای گونه: تجزیه و تحلیل متعارف پرندگان آمریکای جنوبی. J. Biogeogr. 2004 ، 31 ، 1819-1827. [ Google Scholar ] [ CrossRef ]
- المو، اچ. Senay، GB; کاپتو، AT; Kovalskyy، V. تنوع تبخیر و تعرق و ارتباط آن با پویایی پوشش گیاهی در حوضه نیل، 2002-2011. Remote Sens. 2014 , 6 , 5885–5908. [ Google Scholar ] [ CrossRef ]
- توماس، A. ویژگی های مکانی و زمانی روند تبخیر و تعرق بالقوه در چین. بین المللی جی.کلیماتول. 2000 ، 20 ، 381-396. [ Google Scholar ] [ CrossRef ]
- Currie، DJ Energy و الگوهای در مقیاس بزرگ غنای گونه های حیوانی و گیاهی. صبح. نات 1991 ، 137 ، 27-49. [ Google Scholar ] [ CrossRef ]
- روجیرو، آ. Kitzberger، T. همبستگی های زیست محیطی غنای گونه های پستانداران در آمریکای جنوبی: اثرات ساختار فضایی، طبقه بندی و محدوده جغرافیایی. اکوگرافی 2004 ، 27 ، 401-417. [ Google Scholar ] [ CrossRef ]
- مو، کیو. ژائو، ام. در حال اجرا، بهبود SW به الگوریتم جهانی تبخیر و تعرق زمینی MODIS. Remote Sens. Env. 2011 ، 115 ، 1781-1800. [ Google Scholar ] [ CrossRef ]
- لیو، ز. شائو، کیو. لیو، جی. عملکرد محصولات MODIS-GPP و-ET در چین و حساسیت آنها به داده های ورودی (fPAR/LAI). Remote Sens. 2014 ، 7 ، 135-152. [ Google Scholar ] [ CrossRef ]
- جنکینز، CN; Pimm، SL; Joppa، LN الگوهای جهانی تنوع و حفاظت از مهره داران زمینی. Proc. Natl. آکادمی علمی ایالات متحده آمریکا 2013 ، 110 ، 2602–2610. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Pimm، SL; جنکینز، CN; آبل، آر. بروکس، TM; گیتلمن، جی ال. Joppa، LN; ریون، PH; رابرتز، سی ام. Sexton، JO تنوع زیستی گونه ها و میزان انقراض، توزیع و حفاظت از آنها. علم 2014 . [ Google Scholar ] [ CrossRef ] [ PubMed ]
- ژانگ، سی. کای، دی. گوا، اس. گوان، ی. فرادریش، ک. نی، ی. لیو، ایکس. Bian، X. پویایی فضایی-زمانی تنوع زیستی زمینی چین: تشخیصی شاخص زیستگاه پویا. Remote Sens. 2016 , 8 , 227. [ Google Scholar ] [ CrossRef ]
- Parrott, L. اندازه گیری پیچیدگی اکولوژیکی. Ecol. اندیک. 2010 ، 10 ، 1069-1076. [ Google Scholar ] [ CrossRef ]
- مروان، ن. رومانو، ام سی؛ تیل، م. کرتز، جی. نمودارهای بازگشتی برای تجزیه و تحلیل سیستم های پیچیده. فیزیک Rep. 2007 , 438 , 237-329. [ Google Scholar ] [ CrossRef ]
- مروان، ن. وسل، ن. مایرفلد، یو. Schirdewan، A. کورتس، جی. معیارهای پیچیدگی مبتنی بر نمودار عود و کاربرد آنها برای دادههای تغییرپذیری ضربان قلب. فیزیک Rev. E 2002 , 66 , 026702. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- رائو، CR; توتنبورگ، اچ. شلاب، HC; Schomaker, M. مدل های خطی و تعمیم. حداقل مربعات و جایگزین ها ، ویرایش سوم. Springer: برلین، آلمان، 2008. [ Google Scholar ]
- ابربانل، HD; براون، آر. سیدورویچ، جی جی. Tsimring، LS تجزیه و تحلیل داده های آشفته مشاهده شده در سیستم های فیزیکی. Rev. Mod. فیزیک 1993 ، 65 ، 1331. [ Google Scholar ] [ CrossRef ]
- Cao, L. روش عملی برای تعیین حداقل بعد تعبیه یک سری زمانی اسکالر. Physica D 1997 ، 110 ، 43-50. [ Google Scholar ] [ CrossRef ]
- فریزر، AM; Swinney، HL Independent از اطلاعات متقابل برای جذب جاذبه های عجیب و غریب هماهنگ می کند. فیزیک Rev. A 1986 , 33 , 1134. [ Google Scholar ] [ CrossRef ]
- کیان، اچ. Kisling، WD مقیاس فضایی و تطابق متقابل تاکسونی مهرهداران زمینی و گونههای گیاهی آوندی در چین. اکولوژی 2010 ، 91 ، 1172-1183. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- کر، جی تی؛ Packer، L. ناهمگونی زیستگاه به عنوان یک عامل تعیین کننده غنای گونه های پستانداران در مناطق پر انرژی. طبیعت 1997 ، 385 ، 252-254. [ Google Scholar ] [ CrossRef ]
- کیان، اچ. وانگ، ایکس. وانگ، اس. لی، ی. عوامل تعیین کننده محیطی غنای گونه های دوزیستان و خزندگان در چین. اکوگرافی 2007 ، 30 ، 471-482. [ Google Scholar ] [ CrossRef ]
- هاوکینز، کارشناسی; مک کین، سی ام. دیویس، تی جی; باکلی، LB; Anacker، BL; کرنل، HV; دامشن، EI; گریتنس، جی. هریسون، اس. Holt، RD تاریخچه های مختلف تکاملی زمینه ساز شیب غنای گونه های متجانس پرندگان و پستانداران است. J. Biogeogr. 2012 ، 39 ، 825-841. [ Google Scholar ] [ CrossRef ]
- ژائو، ز. لیو، جی. پنگ، جی. لی، اس. وانگ، ی. ویژگیهای غیرخطی و الگوهای پیچیدگی پویایی پوشش گیاهی در منطقه گذار شمال چین. Ecol. اندیک. 2015 ، 49 ، 237-246. [ Google Scholar ] [ CrossRef ]
- Fjeldså، J. بووی، RCK؛ راهبک، ج. نقش رشتهکوهها در تنوعبخشی پرندگان. آنو. کشیش اکول. تکامل. سیستم 2012 ، 43 ، 249-265. [ Google Scholar ] [ CrossRef ]
- هورتال، جی. Carrascal، LM; تریانتیس، کالیفرنیا؛ Thébault، E. میری، س. Sfenthourakis، S. غنای گونه ای می تواند با ارتفاع کاهش یابد اما با تنوع زیستگاهی کاهش نمی یابد. Proc. Natl. آکادمی علمی ایالات متحده آمریکا 2013 ، 110 ، 2149-2150. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- تیوس، جی. بروس، یو. گریم، وی. تیلبورگر، ک. ویچمن، ام سی؛ شواگر، ام. Jeltsch، F. تنوع گونه های جانوری ناشی از ناهمگونی/تنوع زیستگاه: اهمیت ساختارهای کیستون. J. Biogeogr. 2004 ، 31 ، 79-92. [ Google Scholar ] [ CrossRef ]
- کوریستین، LE; کر، JT تغییرات جغرافیایی مرتبط با دما در میان رهگذران: فرآیندهای متضاد در امتداد حاشیههای محدوده قطب و استوا. Ecol. تکامل. 2015 ، 5 ، 5162-5176. [ Google Scholar ] [ CrossRef ]
- توماس، سی دی آب و هوا، تغییرات آب و هوا و محدوده محدوده. غواصان. توزیع کنید. 2010 ، 16 ، 488-495. [ Google Scholar ] [ CrossRef ]
- Thuiller, W. Biodiversity: تغییر آب و هوا و اکولوژیست. طبیعت 2007 ، 448 ، 550-552. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- کای، دی. فرادریش، ک. سیلمان، اف. ژانگ، ال. زو، ایکس. گوا، اس. Guan، Y. پویایی گیاهی در فلات تبت (1982 تا 2006): یک انتساب توسط تشخیص اکو هیدرولوژیکی. جی. کلیم. 2015 ، 4576-4584. [ Google Scholar ] [ CrossRef ]
- چن، اچ. زو، س. پنگ، سی. وو، ن. وانگ، ی. نیش، ایکس. گائو، ی. زو، دی. یانگ، جی. تیان، جی. و همکاران اثرات تغییرات آب و هوایی و فعالیت های انسانی بر چرخه های بیوژئوشیمیایی در فلات چینگهای-تبت گلوب. چانگ. Biol. 2013 ، 19 ، 2940-2955. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- وانگ، ام. ژو، سی. وو، L. الگوی خشکی فلات تبت و عوامل تأثیرگذار آن در 2001-2010. Adv. صعود چانگ. Res. 2012 ، 8 ، 320-326. [ Google Scholar ]
- فن، ال. لو، سی. یانگ، بی. چن، Z. روند بلند مدت بارش در دشت چین شمالی. جی. جئوگر. علمی 2012 ، 22 ، 989-1001. [ Google Scholar ] [ CrossRef ]
- منگ، اس. فی، ی. ژانگ، ز. لی، تی. کیان، ی. Li، Y. تحقیق در مورد توزیع مکانی و زمانی مقدار نفوذ بارش در 50 سال گذشته در دشت شمال چین. Adv. علوم زمین 2013 ، 8 ، 9. [ Google Scholar ]
- اوچیدا، ای. خو، جی. Rozelle، S. Grain برای سبز: مقرون به صرفه بودن و پایداری برنامه کنار گذاشته حفاظت از چین. اقتصاد زمین 2005 ، 81 ، 247-264. [ Google Scholar ] [ CrossRef ]
- خو، ز. خو، جی. دنگ، ایکس. هوانگ، جی. اوچیدا، ای. Rozelle، S. Grain برای سبز در مقابل غلات: تضاد بین امنیت غذایی و حفاظت در چین کنار گذاشته شده است. توسعه دهنده جهانی 2006 ، 34 ، 130-148. [ Google Scholar ] [ CrossRef ]
- چن، ی. وانگ، ک. لین، ی. شی، دبلیو. آهنگ، ی. او، X. متعادل کردن تجارت سبز و غلات. نات Geosci. 2015 ، 8 ، 739-741. [ Google Scholar ] [ CrossRef ]
- لی، ز. ژنگ، فلوریدا؛ لیو، WZ; Flanagan، DC توزیع فضایی و روندهای زمانی رویدادهای شدید دما و بارش در فلات لس چین طی سالهای 1961-2007. کواترن. بین المللی 2010 ، 226 ، 92-100. [ Google Scholar ] [ CrossRef ]
- وانگ، کیو. فن، X. وانگ، ام. روند بارش در طول 1961-2010 در منطقه فلات لس چین. Acta Ecol. گناه 2011 ، 31 ، 5512-5523. [ Google Scholar ]
- چی، تی. ژانگ، Q. وانگ، ی. شیائو، ام. لیو، جی. Sun، P. الگوهای فضایی-زمانی تبخیر تشت در 1960-2005 در چین: تغییر خواص و علل احتمالی. علمی جغرافیا. گناه 2015 ، 35 ، 1599-1606. [ Google Scholar ]
- لیو، جی. کوانگ، دبلیو. ژانگ، ز. خو، X. Qin، Y. نینگ، جی. ژو، دبلیو. ژانگ، اس. لی، آر. Yan, C. ویژگیهای فضایی و زمانی، الگوها و علل تغییرات کاربری زمین در چین از اواخر دهه 1980. جی. جئوگر. علمی 2014 ، 24 ، 195-210. [ Google Scholar ] [ CrossRef ]
- او، تی. Shao، Q. تغییرات مکانی-زمانی تبخیر و تعرق زمینی در چین از سال 2001 تا 2010 با استفاده از محصولات MOD16. J. Geo-Inform. Sci 2014 , 16 , 979-988. [ Google Scholar ]
- میدن، RL سلسله مراتبی از مفاهیم گونه: انحراف در حماسه مشکل گونه. در گونه: واحدهای تنوع زیستی ; Claridge, MF, Dawah, HA, Wilson, MR, Eds. چپمن و هال: لندن، انگلستان، 1997; جلد 54، ص 381–423. [ Google Scholar ]
- جیانگ، ز. ممکن است.؛ وو، ی. وانگ، ی. ژو، ک. لیو، اس. فنگ، ز. Li، L. تنوع پستانداران چین و توزیع جغرافیایی ; China Science Publishing: Beijing, China, 2015. (به زبان چینی) [ Google Scholar ]
- هبرت، PD; Cywinska، A.; شناسایی بیولوژیکی توپ، SL از طریق بارکدهای DNA. Proc. R Soc. Lond B Bio. علمی 2003 ، 270 ، 313-321. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- فریتز، اس. مک کالوم، آی. شیل، سی. پرگر، سی. ببینید، L. شپاچنکو، دی. ون در ولده، م. کراکسنر، اف. Obersteiner, M. Geo-wiki: یک پلتفرم آنلاین برای بهبود پوشش جهانی زمین. محیط زیست مدل. نرم افزار 2012 ، 31 ، 110-123. [ Google Scholar ] [ CrossRef ]


© 2017 توسط نویسندگان. دارنده مجوز MDPI، بازل، سوئیس. این مقاله یک مقاله با دسترسی آزاد است که تحت شرایط و ضوابط مجوز Creative Commons Attribution (CC BY) ( http://creativecommons.org/licenses/by/4.0/ ) توزیع شده است.
بدون نظر