

خلاصه
زنبورداری برای تولید جهانی غذا ضروری است. این یک منبع درآمد جایگزین، به ویژه در سکونتگاه های توسعه نیافته آفریقایی روستایی، و یک انگیزه مهم حفاظت از جنگل است. با این حال، کاهش مستعمرات زنبور عسل در سراسر جهان به آفات و بیماری هایی نسبت داده می شود که توزیع مکانی و تأثیرات آنها به خوبی مشخص نشده است. در این مطالعه، ما از دادههای سنجش از دور برای بهبود قابلیت اطمینان مدلهای طاقچه اکولوژیکی آفات (EN) برای دستیابی به نقشههای توزیع آفات قابل اعتماد استفاده کردیم. داده های بروز چهار آفت ( Aethina tumida , Galleria mellonella , Oplostomus haroldi و Varroa destructor )) از زنبورستان های چهار منطقه اصلی کشاورزی-اکولوژیکی مسئول بیش از 80 درصد از زنبورداری کنیا جمع آوری شد. متغيرهاي زيست اقليمي Africlim و شاخص گياهي تفاوت نرمال شده مشتق شده (NDVI) براي مدلسازي طاقچه هاي اكولوژيكي آنها با استفاده از حداكثر آنتروپي (MaxEnt) استفاده شد. متغیرهای بارش ترکیبی تأثیر لاجیت مثبت بالایی بر عملکرد همه مدلهای سنجش از دور و زیستی داشتند. متغیرهای پوشش گیاهی سنجش از دور تأثیر قابلتوجهی بر مدل داشتند و تا 40.8 درصد برای G. mellonella مشارکت داشتند.و مناطقی با فصلی بودن بارندگی بالا به عنوان مناطق پرخطر پیشبینی شد. پیشبینیها (تا سال 2055) نشان داد که با روند تغییرات آب و هوایی فعلی، این مناطق با افزایش خطر آفت زنبور عسل مواجه خواهند شد. نتیجه میگیریم که آفات زنبور عسل را میتوان با استفاده از دادههای زیستاقلیمی و متغیرهای سنجش از دور در MaxEnt مدلسازی کرد. اگرچه دادههای زیستاقلیمی در همه نتایج مدل بسیار مرتبط بودند، ترکیب متغیرهای فصلی پوشش گیاهی برای بهبود نقشهبرداری از زیستگاه «واقعی» آفات کلیدی زنبور عسل و شناسایی مناطق خطر و مهار باید بیشتر مورد بررسی قرار گیرد.
کلید واژه ها:
آفات زنبور عسل ; متغیرهای زیست اقلیم ; متغیرهای سنجش از دور ; فنولوژی ; مدل سازی طاقچه اکولوژیکی
چکیده گرافیکی
1. معرفی
زنبورداری یک فعالیت اقتصادی مهم در سطح جهان است. خدمات گرده افشان آن به ویژه برای تولید جهانی غذا و امنیت غذایی ضروری است [ 1 ، 2 ]. علاوه بر این، زنبورداری به حفاظت از منابع طبیعی، به ویژه در جوامعی که در اطراف جنگل ها زندگی می کنند، کمک می کند. این امر درآمد خانوارها را در دشت های آفریقایی که اغلب تحت سلطه بارندگی نامنظم و غیرقابل اعتماد است که قادر به حمایت کافی از کشاورزی دیم نیست، متنوع می کند [ 3]]. درآمد اضافی حاصل از فروش محصولات کندو مانند عسل، موم، بره موم، ژل رویال و تا حدی زهر زنبور عسل است. با این حال، سلامت زنبور عسل، خدمات گرده افشانی و مزایای معیشتی مرتبط با تغییرات آب و هوایی، تغییر زیستگاه (تجزیه و از بین رفتن)، تشدید کشاورزی، وابستگی بیش از حد به مواد شیمیایی کشاورزی، و به طور فزاینده ای توسط عوامل بیماری زا، آفات و بیماری ها تهدید می شود [4 ، 5 ] .
آفات به خصوص به دلیل پوشش فضایی و توانایی آنها در ایجاد آسیب مستقیم (از طریق آسیب فیزیکی) و غیرمستقیم (به عنوان ناقل بیماری و بیماری) از نظر اقتصادی مهم ترین در نظر گرفته می شوند. آفات زنبور عسل علیرغم نقش آنها در تخریب کلونی، وقوع و توزیع، همچنان در مناطق استوایی آفریقا مورد مطالعه قرار نگرفته اند، با تحقیقات محدود به کنیا، مالاوی و آفریقای جنوبی [6 ] . علاوه بر این، دادههای مربوط به تغییرات داخل کشور از یک منطقه کشاورزی-اکولوژیکی به منطقه دیگر کمیاب است. در آفریقا، سوسک کندوی کوچک ( Aethina tumida )، سوسک کندوی بزرگ ( Oplostomus haroldi و Oplostomus fuligineus )، پروانه مومی ( Galeria mellonella و Achroia grisella)و کنه Varroa ( Varroa destructor )، آفات مهمی هستند که قادر به ایجاد خسارت اقتصادی قابل توجهی هستند [ 4 ، 6 ، 7 ، 8 ]. در این میان، کنه Varroa به دلیل توانایی آن در ایجاد آسیب مستقیم از طریق عادت تغذیهای خونخوار و بهطور غیرمستقیم از طریق انتقال فعال ویروسهای زنبور عسل، تنها مخربترین آفت برای زنبور عسل است [4 ، 6 ، 9 ] .
اطلاعات کمی از پراکنش مکانی و زمانی این آفات در این قاره وجود دارد. بروز و پراکنش این آفات تحت تأثیر متغیرهای مختلف زیستی و غیرزیستی است. مانند سایر آفات، آفات زنبور عسل می توانند در برخی شرایط بهینه زیست اقلیمی زنده بمانند. به عنوان مثال، دمای بهینه، رطوبت، بارندگی، ارتفاع و محدوده بهره وری اولیه زیست توده/خالص برای آفات مختلف زنبور عسل می تواند به طور قابل توجهی متفاوت باشد [ 10 ]. مطالعات نشان داده اند که توانایی تولیدمثلی آفات زنبور عسل را می توان با شرایط خشک غالب محدود کرد و با شرایط گرم و مرطوب افزایش داد [ 8 ، 11] .]. فنولوژی پوشش گیاهی همچنین می تواند بر پویایی جمعیت آفات زنبور عسل و غنای گونه ای آنها تأثیر بگذارد. فنولوژی گیاهی در دسترس بودن منابع غذایی را تعیین می کند که میزبانان آنها (زنبورهای عسل) از آن برای تولید محصولات کندو مانند عسل و نان زنبور که آفات در آنها رشد می کنند استفاده می کنند. علاوه بر این، رزینهای گیاهی جمعآوریشده توسط زنبورهای عسل برای ساخت کندو و دفاع در برابر ارگانیسمهای خارجی (کپسولاسیون اجتماعی) و خوددرمانی مفید هستند [ 12 ، 13 ]. بنابراین، فنولوژی پوشش گیاهی به طور مستقیم و غیرمستقیم با تأثیرگذاری بر مراحل زندگی آفاتی مانند سوسکهای کندو که در خارج از محیط کندو رخ میدهند، نقش اصلی را در سلامت کلنی ایفا میکند [ 6 ، 14 ].
رویکردهای مدلسازی طاقچه اکولوژیکی (EN) میتواند مسیری را فراهم کند که از نظر آماری تغییرات فضایی در متغیرهای زیستاقلیمی را به توزیع یک گونه خاص (از جمله آفات زنبور عسل) مرتبط کند. مدلهای EN بر همبستگی بین زیستگاههایی که ارگانیسمهای مختلف (مانند آفات زنبور عسل) رشد میکنند و متغیرهای محیطی مانند شرایط زیستاقلیمی، زمین و پوشش گیاهی [ 15 ، 16 ] تکیه میکنند. با این حال، دقت مکان و توزیع زمانی آفات زنبور عسل و زیستگاه آنها عوامل مهمی برای استخراج مدل های دقیق است [ 17 ، 18] .]. مدلهای EN مناسب که میتوانند توزیع فضایی گونههای آفات زنبور عسل را بهطور دقیق پیشبینی کنند، بر دادههای حضور آفت نماینده و دقیق همراه با متغیرهای پیشبینیکننده اکولوژیکی و اقلیمی با دقت انتخاب شده تکیه میکنند [19 ] .
توزیع جغرافیایی دقیق و واقعی گونههای آفات زنبور عسل با استفاده از متغیرهای سنجش از راه دور مانند پویایی “سبز بودن” پوشش گیاهی به دست میآید [ 20 ]. این به این دلیل است که متغیرهای سنجش از دور مشاهدات پیوسته با وضوح مکانی و زمانی بالا در مقایسه با متغیرهای زیست اقلیمی هستند که مدلسازی یا درونیابی میشوند [ 14 ]. داده های سنجش از دور خوب پردازش شده همچنین به کاهش بیش از حد برازش (پیش بینی بیش از حد) مدل های EN کمک می کند و دقت و ارزش گونه ها و برآوردهای توزیع آفات را بهبود می بخشد [20 ] .
اگرچه تعدادی از مطالعات تأثیر عوامل مختلف بیولوژیکی، اقلیمی، پوشش گیاهی و ادافیک را بر برخی از آفات زنبور عسل، مانند سوسکهای کندوی کوچک [21] و سوسکهای کندوی بزرگ [ 7 ، 8] روشن کردهاند.]، سهم فنولوژی پوشش گیاهی و فعل و انفعالات زیست اقلیم در توزیع و فراوانی آنها در کنیا به حداقل مورد بهره برداری قرار می گیرد. بنابراین، نقشه برداری از توزیع فضایی آفات زنبور عسل برای ارزیابی ریسک و پهنه بندی مناطق خطر آفات زنبور عسل برای ایجاد مناطق مهار و مسیرهای انتقال منبع ارزشمند است. تکنیکهای سنجش از دور پیشرفته را میتوان برای استخراج متغیرهای فنولوژیکی (مانند شروع و پایان فصل) از دادههای سنجش از دور که اطلاعات دقیق و مشخصی را برای پیشبینی گسترش آفات زنبور عسل ارائه میدهد، استفاده کرد. مدلی که متغیرهای سنجش از دور و زیست اقلیمی را به هم پیوند می دهد، یک سیستم عملیاتی برای نظارت بر تغییرات بلندمدت مربوط به گسترش آفات ارائه می دهد [ 20]]. همچنین ابزاری ارائه میدهد که میتواند سناریوهای آیندهنگر توزیع آفات زنبور عسل را مدلسازی کند، زیرا آب و هوای پیشبینیشده و پاسخهای فنولوژیکی را میتوان مدلسازی کرد.
در این مطالعه، ما با هدف شناسایی متغیرهای کلیدی اکولوژیکی / سنجش از دور و زیست اقلیمی مرتبط با تکثیر آفات زنبور عسل در کنیا انجام شد. ما همچنین به دنبال ادغام متغیرهای سنجش از دور برای نقشه های توزیع منسجم تر با استفاده از پارامترهای فنولوژی گیاهی مشتق شده بودیم. ما به سناریوهای آینده توزیع این آفات با استفاده از داده های پیش بینی مدل شده اشاره کردیم.
2. روش ها
2.1. سایت های مطالعه
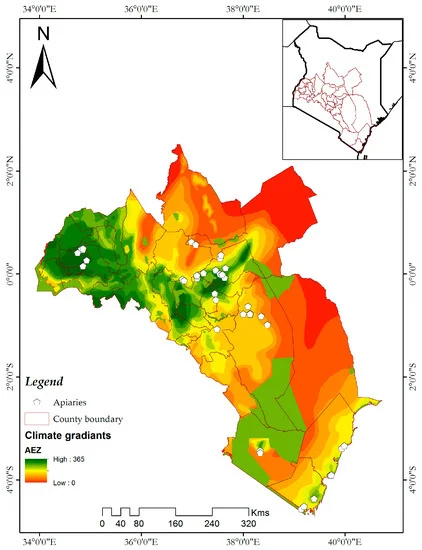
داده های وقوع مورد استفاده در این مطالعه از چهار منطقه اصلی (مناطق کشاورزی-اکولوژیکی) در کنیا جمع آوری شد. ساحل، کوه کنیا، مووینگی و کاکامگا ( شکل 1 )، که بر اساس یک شیب آب و هوایی معرف طبقه بندی می شوند. منطقه ای به وسعت 251322.66 کیلومتر مربع، که از منطقه ساحلی از طریق بخش مرکزی و شرقی کنیا تا دریاچه ویکتوریا را در بر می گیرد، برای مدل سازی سوله های اکولوژیکی این آفات زنبور عسل استفاده شد (شکل 1 ) . این سایت شامل 32 شهرستان اداری است. دما و رطوبت در منطقه ساحلی بالاترین میزان است در حالی که سایت Mwingi در یک منطقه نیمه خشک ساوانا قرار دارد که گرم و خشک است [ 22 ]. کوه کنیا و کاکامگا مناطق مرتفعی هستند که شرایط آب و هوایی خنکتر و مرطوبتری دارند.23 ].
2.2. داده های وقوع
داده های وقوع گونه های آفت زنبور عسل در طول فصل مرطوب (مارس تا آوریل 2014) و فصل خشک (ژوئن 2015) جمع آوری شد. یک زنبورستان با 5-10 کلنی در یک مکان به عنوان یک نقطه نمونه در نظر گرفته شد. چهار گونه آفت زنبور عسل ( A. tumida ، G. mellonella ، O. haroldi و V. destructor ) برای این مطالعه انتخاب شدند. این چهار آفت بیشترین برخورد را دارند و از نظر اقتصادی مهمترین آفات با شیوع گسترده به عنوان آفات مهاجم در سطح جهان هستند [ 7 ، 8 ]. سرشماری آفات در پنج کلنی در هر زنبورستان انجام شد و حضور آنها با استفاده از روشهای استاندارد که توسط Dietemann و همکارانش توضیح داده شد، ثبت شد. [ 24 ] برایA. tumida ، G. mellonella و V. destructor ، و توسط Torto و همکاران. [ 8 ] برای O. haroldi . ما از 37 زنبورستان که دارای A. tumida بودند ، 38 دارای G. mellonella ، 24 دارای O. haroldi و 45 دارای V. destructor در تمام مناطق مورد مطالعه بودند. اینها در اندازه نمونه قابل قبول در محیط مدلسازی MaxEnt بودند [ 10 ، 25 ].
ما فرض کردیم که افراد این گونه ها به طور یکنواخت در سراسر فضا توزیع شده اند. داده های حضور از سایت های مختلف مستقل از متغیرهای محیطی با توجه به احتمال ناشناخته توزیع جمع آوری شد. بنابراین، زنبورستانها بهطور تصادفی با توجه به الزامات تراکم تخمینی نمونهبرداری شدند، که موظف است افراد بهطور تصادفی در سراسر چشمانداز متناسب با تراکم جمعیت نمونهبرداری شوند [ 26 ].
برای آزمایش تأثیر فصلی بر فراوانی آفات زنبور عسل، آزمون U Mann-Whitney برای آزمایش معنیداری ( 05 /0p≤) تفاوت بین فصلهای مرطوب و خشک به دنبال مشاهده واریانس ناهمگن انجام شد.
2.3. آماده سازی داده ها برای تجزیه و تحلیل
تحلیلهای پیشبینی بر اساس مجموعههای مختلف دادههای جغرافیایی، که شطرنجی و برداری بودند، بر روی مرز محل مطالعه برش داده شدند. این مجموعه داده ها به دو گروه اصلی تقسیم شدند. متغیرهای وقوع و پیش بینی کننده متغیرهای پیش بینی کننده به متغیرهای سنجش از دور و بیوکلیماتیک طبقه بندی شدند ( جدول 1 ).
2.4. پردازش داده های سنجش از راه دور
2.4.1. متغیرهای بیوتیک
متغیرهای سنجش از راه دور در دینامیک بهرهوری پوشش گیاهی از دادههای سری زمانی طیفسنج تصویربرداری با وضوح متوسط (MODIS)، در وضوح فضایی 250 متر، پردازش شده و بهعنوان متغیرهای پیشبینیکننده اکولوژیکی در مدلسازی EN (جدول 1) مورد استفاده قرار گرفتند . کامپوزیت های اصلاح شده NDVI 16 روزه MODIS برای سال های 2001 تا 2014 (دوره مشاهده 14 ساله) به دست آمد. مجموعه داده های سری زمانی تصحیح شده بیشتر در نرم افزار TIMESAT [ 27 ، 28 ] برای استخراج متغیرهای فصلی پوشش گیاهی که بخشی از متغیرهای محیطی ورودی در الگوریتم مدل سازی حداکثر آنتروپی را تشکیل می دادند، پردازش شدند ( جدول 1 ).
TIMESAT سیگنالهای فنولوژیکی موجود در دادههای ماهوارهای سری زمانی را با برازش توابع محلی به نقاط داده سری زمانی تجزیه و تحلیل میکند، سپس آنها را در یک مدل جهانی ترکیب میکند [ 29 ، 30 ]. پس از آن، یک تابع مدل صاف برای استخراج متغیرهای فنولوژیکی برای هر فصل رشد استفاده می شود، که به نوبه خود تأثیر نویز سیگنال باقیمانده را در داده های سری زمانی NDVI [ 31 ، 32 ] و ابعاد داده ها [ 33 ، 34] کاهش می دهد.]. پارامترهای برازش عملکرد مورد استفاده در TIMESAT برای این مطالعه عبارت بودند از: یک روش فیلتر ساویتزکی-گولای، پنجره 3 و 4 نقطه ای بیش از 2 مرحله اتصال، قدرت تطبیق 3.0، بدون برش سنبله یا دامنه، قطع فصل 0.0، و شروع و آستانه پایان فصل 20٪.
در مجموع، 12 متغیر سنجش از دور برای هر فصل رشد استخراج شد. زمان شروع فصل ( seas_start ) که 20% از حداقل لبه سمت چپ، زمان پایان فصل ( seas_end ) 20% از حداقل سمت راست، طول فصل یا زمان از شروع تعیین شده است. تا پایان فصل رشد ( seas_length )، سطح پایه، که با میانگین کردن مقادیر حداقل چپ و راست ( سطح_پایه )، زمان برای اواسط فصل ( seas_mid )، بزرگترین مقدار داده برای تابع برازش شده در طول فصل محاسبه شد. ( حداکثر )، دامنه فصلی ( دامنه )، میزان افزایش در ابتدای فصل (left_der )، میزان کاهش در پایان فصل ( right_der )، انتگرال فصلی بزرگ ( large_int )، انتگرال فصلی کوچک ( small_int ) و تعداد فصل ها در یک سال تقویمی ( num_seas ) ( جدول 1 ). فقط از متغیرهای فصل اول در این مطالعه استفاده شد زیرا دادههای فصل دوم در تمام سالها سازگار نبود.
2.4.2. متغیرهای توپوگرافی
متغیرهای توپوگرافی مختلف؛ ارتفاع، شیب، سایه تپه و جنبه در درجه تشعشع ( جدول 1 ) در مدل EN برای پیش بینی وقوع چهار گونه آفات زنبور عسل گنجانده شد. متغیرهای توپوگرافی از داده های مدل رقومی ارتفاع 90 متری (DEM) از ابزارهای ماموریت توپوگرافی رادار شاتل (SRTM) [ 35 ] مشتق شدند. دادههای DEM برای تناسب با اندازه پیکسل 250 متر و گستره فضایی مجموعه دادههای NDVI 16 روزه MODIS نمونهگیری مجدد شدند.
2.5. داده های زیست اقلیمی
برای شبیهسازی توزیع گونههای آفات زنبور عسل، شرایط آب و هوایی فعلی در وضوح شبکه یک کیلومتری از مجموعه دادههای AfriClim استفاده شد [ 36 ، 37 ، 38 ]. این مجموعه داده شامل شبکههایی از دما، بارندگی و متغیرهای خلاصه زیستاقلیمی مشتق شده است. برای پیشبینی توزیع تحت شبیهسازیهای شرایط آب و هوایی آینده، دادههای کوچکشده سناریوهای مسیرهای تمرکز نماینده، گزارش ارزیابی پنجم (RCPs-AR5) [ 39 ] سال آینده 2055 (میانگین بیش از 2041-2070) استفاده شد.
داده های زیست اقلیم ( جدول 1 ) به متغیرهای دما و رطوبت تقسیم شدند. متغیرهای دما شامل میانگین دمای سالانه ( bio1 )، میانگین دمای روزانه ( bio2 )، همدما ( bio3 )، فصلی بودن دما ( bio4 )، حداکثر دمای گرمترین ماه ( bio5 )، حداقل دمای خنکترین ماه ( bio6 )، سالانه بود. محدوده دما ( bio7 )، میانگین دمای گرمترین چهارم ( bio10 )، میانگین دمای سردترین چهارم ( bio11 )، و تبخیر و تعرق بالقوه ( pet). متغیرهای رطوبت بودند. میانگین بارندگی سالانه ( bio12 )، بارندگی مرطوب ترین ماه ( bio13 )، بارندگی خشک ترین ماه ( bio14 )، فصلی بودن بارندگی ( bio15 )، بارندگی مرطوب ترین سه ماهه ( bio16 )، بارندگی خشک ترین سه ماهه ( bio17 )، سالانه شاخص رطوبت ( mi )، شاخص رطوبت یک چهارم مرطوب ( mimq )، شاخص رطوبت یک چهارم خشک ( miaq )، تعداد ماههای خشک ( dm ) و طول طولانیترین فصل خشک ( llds)). همه 21 متغیر آب و هوایی به اندازه سلول 250 متر و به وسعت فضایی مجموعه دادههای NDVI 16 روزه MODIS نمونهبرداری شدند.
2.6. مدل سازی طاقچه اکولوژیکی
سوله های اکولوژیکی چهار آفت زنبور عسل با استفاده از حداکثر آنتروپی (MaxEnt) در بسته ابزار NMaxEnt نسخه 3.3.3k [ 40 ] مدل سازی شدند. احتمال تخمین برای وقوع گونههای آفت زنبور عسل به مجموعهای از پیکسلها از ویژگیها برای به حداکثر رساندن آنتروپی چگالی تخمین تحت محدودیتهای داده شده پدیدههای متغیر ناشناخته برازش داده شد. سپس از مقادیر این پیکسل ها برای تخمین احتمال توزیع این گونه های آفت استفاده شد. نمونهبرداری تصادفی خودکار از نمونههای شبه غیبت/پسزمینه از مجموعهای از پیکسلها در مرزهای کنیا [ 41 ]، جایی که این گونهها شناسایی نشدهاند [ 42 ] برای به حداکثر رساندن پیشبینی استفاده شد. شبه فقدان احتمال حضور که در غیر این صورت فقط به حضور محدود می شد [43 ]. فرض بر این بود که هر گونه احتمال یکسانی برای قرار گرفتن در هر نقطه از چشم انداز دارد، از این رو هر پیکسل یا محیطی در منظره احتمال یکسانی برای برچسب گذاری به عنوان “پس زمینه” در فضای جغرافیایی و محیطی دارد.
2.7. انتخاب متغیر
برای به حداقل رساندن چند خطی در میان متغیرهای پیشبینیکننده [ 26 ]، ما یک آزمون همبستگی پیرسون را بین تمام پیشبینیکنندههای ارائهشده در جدول 1 انجام دادیم تا ویژگیهای متعامد مناسب برای مدل EN را بدست آوریم. ضریب همبستگی |r| > 0.7 [ 44 ] به عنوان یک شاخص همخطی برای متغیرهایی که به شدت بر مدل EN ما تأثیر میگذارند، تنظیم شد. متغیرهایی که این معیار را داشتند بر اساس تحلیل اهمیت متغیر حذف شدند که برای شناسایی آنهایی که قدرت توضیحی بالایی داشتند انجام شد. اجرای آزمایشی با استفاده از همه متغیرهای زیست اقلیمی و سنجش از دور در جدول 1 برای تجزیه و تحلیل سهم آنها انجام شد ( شکل 2)) به هر گونه آفت قبل از ساختن مدل های طاقچه آنها با استفاده از آزمون jackknife [ 45 ].
خلاصه ای از روش در یک نمودار جریان نشان داده شده است ( شکل 3 ). تعاملات متغیرهای مختلف، دادههای وقوع و انتخاب مدل مناسب به صورت گرافیکی نشان داده شده است.
2.8. تنظیمات مدل
ما از تنظیمات پیشفرض MaxEnt با چند تغییر با هدف دستیابی به پیشبینیهای دقیق و دقیق استفاده کردیم [ 19 ، 26 ]. بیست درصد از سوابق وقوع از هر مدل پنهان شد تا به عنوان دادههای آزمون مستقل استفاده شود. با این حال، تنظیمات پیشفرض مدل ما را با نصب بیش از حد جریمه کرد. بنابراین، ما ترجیح دادیم از ضریب تنظیم سه برای پخش کردن پیشبینیها استفاده کنیم و برای به دست آوردن یک مدل قوی، تعداد دفعات تکرار را روی ده تنظیم کنیم. با توجه به افزایش تکرار، از دانه تصادفی برای انتخاب پارتیشن تست/اجرای تصادفی مختلف برای هر اجرا استفاده شد. ما از “آستانه حضور تمرینی 10 صدکی” استفاده کردیم که 10٪ شدیدترین مشاهدات حضور را به عنوان غایب پیش بینی می کند [ 20 ] برای حذف نقاط پرت از مدل.
مدلسازی تحت شرایط زیست اقلیم، پوشش گیاهی و توپوگرافی فعلی انجام شد. به طور خاص، مدلها تنها با استفاده از پیشبینیکنندههای زیستاقلیمی (مدل 1)، تنها متغیرهای سنجش از دور (مدل 2) و هر دو متغیر زیستاقلیمی و سنجش از دور (مدل 3) ترکیب شدند. این مدل ها در آینده (سال 2055) پیش بینی شدند. ما از شبیهسازیهای اقلیمی آینده از Africlim برای طرح سناریوی آینده استفاده کردیم و به دلیل عدم پیشبینی پوشش گیاهی، پوشش گیاهی و توپوگرافی را بدون تغییر در طول دوره پیشبینی فرض کردیم.
2.9. ارزیابی مدل
ما از سطح مستقل آستانه زیر منحنی (AUC) مدل تحلیل مشخصه عملکرد گیرنده (ROC) [ 19 ] برای ارزیابی عملکرد مدل استفاده کردیم. ناحیه زیر یک منحنی ROC احتمال این را نشان می دهد که وجود (حساسیت) در مقابل عدم وجود (ویژگی) یا نقاط پس زمینه [ 46 ] توسط یک طبقه بندی کننده به درستی مرتب شده اند. مقادیر AUC صفر (0) منطقه وقوع غیرممکن را نشان می دهد در حالی که یک (1) منطقه وقوع بهینه یا ترتیب کامل را نشان می دهد [ 47 ]. ما از Swets استفاده کردیم [ 48] قدرت تبعیض آمیز برای رتبه بندی مدل ها به عنوان; (i) عالی = 0.91-1.00، (ii) خوب = 0.81-0.90، (iii) مناسب = 0.71-0.80، (iv) ضعیف = 0.61-0.70، و (v) شکست = 0.51-0.60. برای ارزیابی سودمندی متغیرهای سنجش از دور در بهبود عملکرد مدل، از مدل صفر [ 49 ] استفاده شد. سی و شش نقطه تولید شده به طور تصادفی از سلول های حضور مشاهده شده برای استخراج مقادیر AUC از هر یک از سه مدل (مدل 3، مدل 1 و مدل 2) استفاده شد. برای ارزیابی تفاوت معنیداری (05/0 ≤ p) بین سه مدل از آزمون U Mann-Whitney استفاده شد. ما بیشتر خطای حذف مدلهای مختلف توسعهیافته را محاسبه کردیم، زیرا مدلهای ما فقط از دادههای حضور استفاده میکردند و محاسبه خطای کمیسیون به دادههای غیبت نیاز دارد [ 50 ].
3. نتایج
3.1. فراوانی آفت زنبور عسل
جدول 2 دادههای فراوانی چهار آفت زنبور عسل را در چهار منطقه (کوه کنیا، مووینگی، کاکامگا و مناطق ساحلی) جمعآوریشده در فصول مرطوب و خشک نشان میدهد. به طور کلی آفات در فصل مرطوب نسبت به فصل خشک بیشتر بود. تفاوت معنی داری در فراوانی آفت بین فصول وجود داشت. علاوه بر این، به نظر می رسد A. tumida و V. destructor نسبت به تغییرات بارش حساس تر هستند زیرا فراوانی آنها تقریباً شش برابر افزایش یافته است.
3.2. مدل های EN
جدول 3 AUC مستقل از آستانه را نشان می دهد که از مدل آنالیز ROC به دست آمده است، که عملکرد “خوب” را برای همه مدل 3 و همه مدل 1 به جز G. mellonella و مدل عادلانه 2 برای همه آفات زنبور عسل نشان می دهد. مدل 1 مقادیر AUC بالاتری را برای همه آفات نسبت به مدل 2 به دست آورد. با این حال، مدل 3 نمرات AUC بالاتری برای A. tumida (0.87)، O. haroldi (0.87) و V. destructor (0.88) نسبت به مدل 1 یا مدل 2 داشت. مقادیر AUC برای G. mellonella مدل 3 (0.80) مانند مدل 1 (0.80) بود. با این حال، نقشه های پیش بینی مدل 3، اضافه برازش کمتر با انحراف استاندارد کم را در تمام مدل های گونه های آفات زنبور عسل نشان داد ( جدول 3 ).
تجزیه و تحلیل مدل صفر نشان داد که بین مدل 2 و مدل 1 و 3 برای همه گونه ها به جز O. haroldi تفاوت معنی داری وجود دارد . با این حال، تفاوت معنی داری بین مدل 1 و مدل 3 وجود نداشت. بنابراین، مدل 2 از مقایسه مدل بعدی حذف شد.
مدل های آفت خطای حذف کم برای مدل 3 نسبت به مدل های دیگر برای همه آفات داشتند (به عنوان درصد برای مدل 3، مدل 1 و مدل 2 فهرست شده اند، A. tumida 16.6، 19.4 و 25.0، G. mellonella 27.7، 33.3 و 38.9 ، O. haroldi 30.5، 33.3 و 38.9 و V. distractor 19.4، 25.0 و 30.6). مدل 3 برای تمام آفات زنبور عسل بهتر از مدل های دیگر عمل کرد.
3.3. سهم متغیر پیش بینی کننده
در تمام مدل 3 برای چهار آفت زنبور عسل، ترکیب متغیرهای زیست اقلیم قویترین تأثیر نسبی را نسبت به متغیرهای سنجش از دور ترکیبی داشتند. متغیرهای زیست اقلیم 88.1 درصد برای O. haroldi ، 83.0 درصد برای A. tumida ، 68.3 درصد برای V. destructor ، و 58.7 درصد برای G. mellonella ( جدول 4 ) سهم داشتند . علاوه بر این، در خوشه زیست اقلیم، متغیرهای بارش تأثیر بیشتری (از 58.0٪ تا 81.0٪) نسبت به متغیرهای دما (از 0.4٪ تا 7.3٪) داشتند. از سوی دیگر، متغیرهای سنجش از دور در مدل 3 (زیستی و توپوگرافی) سهم کلی 17.0 درصد برای A. tumida ، 41.3 درصد برای G. mollenela ، 11.9 درصد برایO. haroldi و 31.7% برای V. destructor. سهم نسبی ترکیبی متغیرهای زیستی سنجش از دور در مدل 3 از 11.4% تا 40.8% متغیر بود ( جدول 4 ). در این خوشه متغیر، حداکثر NDVI در فصل سهم نسبی قابل توجهی (بین 7.7٪ و 29.4٪) به مدل های EN داشت. متغیرهای توپوگرافی سهم نسبتاً کمتری داشتند، کمتر از 4.2٪ و هیچ اثر مدلسازی شدهای بر V. destructor نداشتند.
بر اساس آزمایشهای جکنایف، اهمیت متغیر نسبی (هم زیستاقلیمی و هم سنجش از راه دور) به مدل 3 ( شکل 4 )، سهم متفاوتی را در مدلهای مختلف EN نشان داد. پنج متغیر اول رتبه بندی شده (bio15، mimq، max_ndvi، دامنه و شیب) به طور تجمعی به 94.3٪ در مدل A. tumida کمک کردند . پنج متغیر اول (mimq، max_ndvi، bio12، bio15 و base_level) 96.3% سهم تجمعی در مدل G. mellonella و 92.8% در مدل V. destructor داشتند ، در حالی که bio15، bio3 و max_ndvi سهم تجمعی 95.3% در O داشتند. مدل هارولدی
Bio15 در مدل 3 بیشترین افزایش را در A. tumida (43.9٪)، O. haroldi (79.7٪) و V. destructor ( 28.2 ٪) داشت . ٪. همه این متغیرها در مدل مربوطه 3 دارای لاجیت مثبت بودند. متغیرهای توپوگرافی هیچ تأثیری بر V. destructor نداشتند و تأثیر کمی بر مدلهای G. mellonella و O. haroldi نداشتند. از سوی دیگر، A. tumida و G. mellonella مدل 1 نشان دادند که mimq بیشترین تأثیر مثبت را بر توزیع آنها داشت، در حالی که bio12 بر V. destructor تأثیر گذاشت.اکثر. مدل O. haroldi 1 بیشتر تحت تأثیر bio15 قرار گرفت. در مقایسه با سایر متغیرهای سنجش از دور، max_ndvi بیشترین تأثیر را در هر چهار مدل EN مربوط به چهار آفت زنبور عسل نشان داد.
3.4. تجسم توزیع
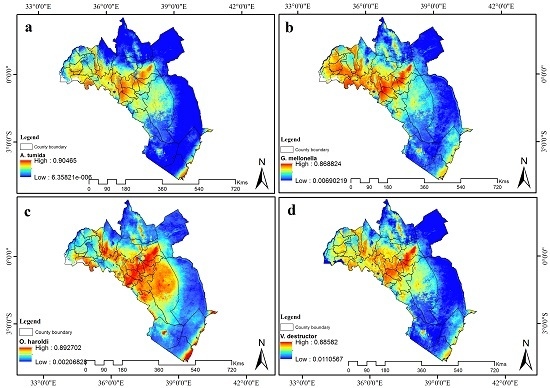
مدلهای آفت زنبور عسل مناسب بودن زیستگاه بالایی را در کوه کنیا و مناطق ساحلی برای هر چهار آفت زنبور عسل پیشبینی کردند ( شکل 5 ). منطقه Mwingi دارای تناسب زیستگاه کم ( G. mellonella ) تا زیاد ( O. haroldi ) پیشبینی شد. با این حال، منطقه کاکامگا احتمال مناسب بودن همه آفات زنبور عسل را داشت. پیشبینی شد که این منطقه دارای خطر O. haroldi کم، ویرانگر V. متوسط تا A. tumida نسبتاً بالا و مناسب زیستگاه G. mellonella بالا باشد ( شکل 6)). کوه کنیا، کاکامگا و مناطق ساحلی با بارش فصلی زیاد مشخص می شوند. با این حال، منطقه ساحلی دما و رطوبت بالاتری نسبت به مناطق کوه کنیا و کاکامگا داشت. از طرف دیگر، مووینگی نسبت به کوه کنیا و کاکامگا دمای نسبتاً بالا و همدما کمتری داشت. اگرچه مناطق ساحلی و کوه کنیا فصلی بارندگی مشابهی داشتند، اما دارای گرادیان دما و همدما متفاوت بودند. در مقابل، منطقه ساحلی و Mwingi میانگین دمای سالانه بالاتر و همدما کمتری نسبت به سایر مناطق داشتند، اما پتانسیلهای خطر آفت زنبور عسل متفاوتی را نشان دادند.
مدلهای سناریوی آیندهنگر برای چهار آفت زنبور عسل به طور کلی نشان داد که مناطق مناسب زیستگاهی بالا ممکن است به مناطقی که در حال حاضر به عنوان دارای تناسب آفت متوسط تا کم طبقهبندی شدهاند گسترش یابد ( شکل 6 ). دادههای زیستاقلیمی مدلسازیشده اساساً افزایش میزان بارندگی در مرطوبترین سهماهه و افزایش تنوع بارندگی را پیشبینی میکنند.
3.5. مشارکت داده های سنجش از راه دور
متغیرهای سنجش از دور دقت و دقت مدل ها را با کاهش بیش از حد برازش و اصلاح قدرت پیش بینی آنها افزایش دادند ( جدول 3 ). زیرمجموعه ای از نقشه های ویرانگر V. نشان داد که مدل 1 دارای توزیع بیش از حد پیش بینی شده در مقایسه با مدل 3 است، که دارای متغیرهای سنجش از راه دور با اندازه پیکسل کوچکتر (رزولیشن بالا) و ناهمگونی محیطی بیشتر است (شکل 7 ) .
مدل 3 دارای میانگین تناسب زیستگاه کمتری نسبت به مدل 1 برای تمام آفات زنبور عسل مورد بررسی و انحراف معیار کمتر برای A. tumida و V. destructor بود ( شکل 8 ). هیستوگرام مدل 3 نشان می دهد که پیش بینی حول میانگین ادغام شده است در حالی که هیستوگرام bio_model چولگی را به سمت راست نشان می دهد. ادغام در اطراف میانگین توسط متغیرهای سنجش از دور انجام شد، که اندازهگیری آنها در اندازه دانه تضمین میکند که تنوع با دقت بالاتر محدود شده توسط وضوح فضایی قابل تشخیص است. علاوه بر این، مدل 3 مقادیر AUC رتبهبندی بالاتر و انحراف استاندارد کمتری نسبت به مدل 1 برای همه مدلها داشت به جز G. mellonalla ، که با این حال، انحراف استاندارد کمتری نسبت به مدل 1 داشت ( جدول 3) .).
4. بحث
درک عواملی که بر تکثیر و انتشار آفات زنبور عسل در ابعاد مختلف فضایی و سطح خطری که آنها ایجاد می کنند در سلامت زنبور عسل مهم است. بنابراین، پیشبینیپذیری بهبود یافته تنوع آفات و نقشههای توزیع دقیق آفات زنبور عسل ابزار و اطلاعات لازم برای کنترل آفات زنبور عسل را فراهم میکند [ 51 ]. این مطالعه به دنبال شناسایی شرایطی بود که باعث افزایش آفات زنبور عسل در کنیا میشود و مدلهایی را توسعه داد که میتوان از آنها برای پیشبینی گسترش آنها استفاده کرد. مدل های پیش بینی شده برای نشان دادن سطح خطر ناشی از آفت زنبور عسل برای زنبورداران در آینده خدمت کردند.
دادههای نظارتی ما وجود تنوع در فراوانی و توزیع چهار گونه آفت زنبور عسل را در سایتهای مورد مطالعه تأیید کرد. رکوردهای بیشتری در طول فصل مرطوب نسبت به فصل خشک مشاهده شد. این یافته ها با مطالعه قبلی تورتو و همکاران مطابقت داشت. [ 8 ] که تعداد سوسکهای کندو را در هنگام بارندگی بیشتر از فصول خشک کنیا نشان داد. با این حال، تغییرات فصلی مشاهدهشده در جمعیت کنههای Varroa برخلاف گزارشهای قبلی از آلودگی نسبتاً ثابت کنه در سراسر Apis mellifera scutellata (زنبور عسل ساوانا) است [ 52 ]. علاوه بر این، مناطق پست ساحلی توسط A. m. litoreaو ارتفاعات جنگلی توسط زنبورهای مونتیکولا مانند، که با زنبورهای ساوانا در طول مسیرهای مهاجرت خود هیبرید می شوند [ 53 ]. بنابراین، نیاز به درک بیشتر تعامل این آفات زنبور عسل در شکاف های مختلف فضایی وجود دارد.
4.1. سهم متغیر پیش بینی کننده
اگرچه اکثر آفات زنبور عسل دارای محدوده فضایی وسیع تری هستند و می توان آنها را در مناطق مختلف با شرایط آب و هوایی مختلف یافت [ 54 ]، اما شرایط اقلیمی خاصی وجود دارد که نقش کلیدی در تعیین پراکنش آنها دارد. به عنوان مثال، سوسک های کندو و پروانه مومی می توانند در منابع غذایی جایگزین رشد کنند در حالی که واروآیک انگل خارجی اجباری است. بنابراین، دو مورد اول بیشتر از دومی تحت تأثیر تغییرات آب و هوایی قرار دارند. بنابراین ما چهار متغیر کلیدی زیست اقلیمی و سنجش از دور را شناسایی کردیم که مهمترین آنها در تعیین وقوع چهار آفت زنبور عسل هستند. فصلی بودن بارندگی، میانگین بارندگی سالانه، شاخص رطوبت در سه ماهه مرطوب و بیشترین مقدار NDVI در فصل بیشترین تأثیر منطقی را بر مدل 3 برای چهار آفت زنبور عسل داشتند (جدول 4 ) .
این متغیرها عمدتاً با بارش جدا از حداکثر NDVI در فصل مرتبط هستند. علاوه بر این، دادههای وقوع مزرعه ( جدول 2 ) نشان داد که تعداد آفات بیشتری در فصل مرطوب نسبت به فصل خشک ثبت شده است. نتایج مشابهی توسط Torto و همکاران ارائه شد. [ 8 ] که مستند کرد که اثر مثبت بارندگی بر واروآ به طور غیرمستقیم با تولید علوفه زنبور عسل که بر رشد کلنی ها و افزایش تولید نسل تأثیر می گذارد، مرتبط است. این به نوبه خود مکان های تولید مثلی زیادی را برای کنه فراهم می کند [ 24]. نقشه های پراکنش به تأثیر بارندگی در بروز و پراکنش آفت زنبور عسل اشاره می کند. وقوع و پراکنش در مناطقی مانند کوه کنیا، کاکامگا و نوار ساحلی زیاد بود. بنابراین، این چهار متغیر بارش را می توان برای پیش بینی موفقیت آمیز تغییرات فصلی این آفات زنبور عسل در مناطق مختلف داخل کشور استفاده کرد [ 8 ]. از آنجایی که «بارش» یک متغیر بسیار مهم است و با توجه به افزایشهای آینده در مقادیر و تنوع پیشبینیشده بارش [ 36 ، 37 ]، ممکن است از چهار آفتی که ما بررسی کردیم، رخدادهای بالاتری وجود داشته باشد.
مدلهای EN ما نشان داد که توپوگرافی تأثیر کمی بر وقوع آفات زنبور عسل مدلسازیشده در کنیا دارد. مدلها هر دو منطقه با ارتفاع بالا (مانند کوه کنیا) و ارتفاع کم (مانند ساحل) را برای زیستگاه مناسب پیشبینی کردند. این با نتیجه مانی [ 55 ] مطابقت داشت که اکثر حشرات (از جمله آفات زنبور عسل) دارای شیب ارتفاعی متنوعی هستند. در واقع، آفت زنبور عسل که ما بررسی کردیم در تمام شیب های ارتفاعی مناسب زیستگاه یکسانی داشت.
4.2. مشارکت داده های سنجش از راه دور
گنجاندن متغیرهای سنجش از دور در مدلهای EN ما توانایی آنها را برای پیشبینی توزیع گونههای آفات زنبور عسل بهبود بخشید. اگرچه متغیرهای زیست اقلیم توانایی پیشبینی توزیع بالقوه آفات زنبور عسل را ارائه میدهند [ 56 ]، متغیرهای سنجش از دور قابلیت پیشبینی مدل 3 را برای چهار آفت با کاهش بیش از حد برازش، همانطور که در شکل 7 نشان داده شده است، افزایش دادند.. توانایی پیشبینی این مدلهای EN بهبود یافت، زیرا متغیرهای سنجش از راه دور شرایط محیطی تکهتکهشده را روی زمین ثبت کردند. وضوح فضایی خوب متغیرهای سنجش از دور، خطاهایی را که از تعمیم متغیرهای زیست اقلیم در گسترههای فضایی معین ناشی میشوند، کاهش داد. علاوه بر این، محصولات مدل (نقشه های پیش بینی) در پیش بینی تناسب زیستگاه در مدل 3 واقع بینانه تر بودند و تخمین بیش از حد را کاهش دادند [ 20 ، 57 ].
متغیرهای سنجش از دور همچنین شرایط محیطی نامطلوب را که توزیع این آفات زنبور عسل را در شرایط زیست اقلیمی همگن محدود میکند، ثبت میکنند. مدل 1 وقوع مشابهی را در مناطقی با شرایط زیست اقلیمی مشابه که در شکل 7 نشان داده شده است، پیش بینی می کند . با این حال، با متغیرهای سنجش از راه دور مانند حداکثر NDVI در فصل، پتانسیل آفت زنبور عسل بالا توسط مدل 3 در مناطقی با مقادیر NDVI بالا در مقایسه با مناطقی که مقادیر کم را ثبت کردند، پیشبینی شد. تورتو و همکاران [ 8 ] نشان داد که تولید مثل آفات زنبور عسل تحت تأثیر فنولوژی گیاهی مانند NDVI است. علاوه بر این، پاو و همکاران. [ 14] همچنین نشان داد که موفقیت آفات زنبور عسل با در دسترس بودن گرده و شهد تعیین می شود که میزبان آنها به آن بستگی دارد. بنابراین، متغیرهای سنجش از دور که این تغییرات را در بر میگیرند (یعنی متغیرهای NDVI) اطلاعات مدل 3 را ارائه میکنند که در مدل 1 موجود نیست. متغیرهای سنجش از دور همچنین توانایی اصلاح پیشبینی توزیع آفات زنبور عسل را بر خلاف متغیرهای زیستاقلیمی که منعکسکننده هستند، ارائه میکنند. توزیع احتمالی آفت زنبور عسل [ 20 ]. علاوه بر این، متغیرهای محیطی سنجش از دور ناهمگونی را که منعکس کننده شرایط زمین بر اساس زمان واقعی یا تقریباً واقعی است، ثبت میکنند، در حالی که دادههای زیستاقلیمی در مناطق بزرگ درونیابی میشوند، بنابراین همگنی را در گسترههای فضایی بزرگ نشان میدهند [58 ، 59 ] .
4.3. مزایای استفاده از مدل های یکپارچه EN و قابلیت کاربرد
مدلهای EN برای تخمین پیشبینیها تنها به وقوع متکی هستند. در دسترس بودن دادههای غیبت، دقت مدل را بهبود میبخشد، اما چنین دادههایی به آسانی در دسترس نیستند و از این رو نمیتوان با قطعیت نتیجه گرفت که عدم وقوع در یک منطقه، عدم وجود را تأیید میکند [ 60 ]. بنابراین، مدلهای EN از شبه عدم وجود برای پر کردن این شکاف استفاده میکنند. با این حال، اطلاعات مهمی مانند فراوانی یک آفت در یک منطقه خاص که می تواند پیش بینی را بهبود بخشد، مورد استفاده قرار نمی گیرد. دادههای مختلف سنجش از دور با وضوحهای فضایی-زمانی و طیفی مختلف در دسترس هستند، بنابراین مرزهای مدلسازی EN را گسترش میدهند [ 20]]. روشهای تحلیلی پیشرفته دادههای سنجش از دور نیز توانایی استخراج و استفاده از اطلاعات فنولوژیکی خاص از دادههای خام را برای بهبود قابلیت تشخیص ارائه میدهند.
اگرچه دادههای سنجش از راه دور قبلاً برای نقشهبرداری از پراکنش گونههای مختلف گیاهی استفاده شده است [ 20 ، 61 ]، مدلهای ما ابزارهایی را ارائه میدهند که میتوانند برای نقشهبرداری دقیق توزیع آفات زنبور عسل و احتمالاً سایر گونههای حشرات مورد استفاده قرار گیرند. این به این دلیل است که دادههای سنجش از راه دور گنجانده شده در مدلهای ما اطلاعات بهموقع (اخیر) را ارائه میدهند که تغییرات پوشش گیاهی را در نتیجه استفاده از زمینهای انسانی، بهویژه در مناطق کشاورزی یا جنگلهای حفاظتنشده تحت بررسی، امکانپذیر میسازد. در نتیجه، ما ابزارهایی را ارائه میدهیم که میتوانند در روشهای مشابه یا طراحی متفاوتی که در اینجا مستند شدهاند، برای بهبود مدیریت آفات استفاده شوند.
با این حال، داده های سنجش از دور دارای کاستی هایی هستند که می تواند بر مدل های پیش بینی تأثیر منفی بگذارد. از آنجایی که اندازه پیکسل یک شاخص مهم برای کیفیت داده ها است، دقت به دست آمده توسط مدل های پیش بینی که از متغیرهای سنجش از راه دور استفاده می کنند به اندازه پیکسل محدود می شود. بنابراین، مدلهای EN که از دادهها با وضوح درشت استفاده میکنند ممکن است پیشبینی دقیقی ارائه نکنند [ 57]. از سوی دیگر، داده های سنجش از راه دور با وضوح فضایی و طیفی خوب، مانند World View-2 و 3، AISA Eagle، پرهزینه و در دسترس نیستند. علاوه بر این، چالشهایی مانند پوشش ابری کیفیت دادههای سنجش از راه دور نوری را تا حد زیادی کاهش میدهد، زیرا ممکن است اندازهگیریهای محیطی بهدقت ثبت نشده یا زمانی که پوشش ابری قابلتوجه است، در دسترس نباشد. اما ممکن است با وضوح زمانی بالا که گزینههایی برای دور زدن دادهها با کیفیت پایین یا استفاده از فناوری سنجش از راه دور مایکروویو که تحت تأثیر پوشش ابری قرار نمیگیرد، برطرف شود.
مدلهای EN به متغیرهایی با اندازه پیکسل یکسان (یعنی وضوح فضایی) در تحلیل نیاز دارند. به ندرت داده های مکانی از منابع مختلف دارای وضوح مکانی مشابه هستند. نمونهبرداری مجدد از مجموعه دادهها برای دستیابی به اندازه پیکسل یکسان میتواند عدم قطعیت [ 62 ، 63 ] را برای مدلهای EN ایجاد کند. چنین عدم قطعیت ها و موارد مرتبط با جمع آوری داده های حضور ممکن است گمراه کننده باشد [ 64]. بنابراین، دادههای سنجش از راه دور مورد استفاده برای ساخت مدلهای پیشبینی باید با دقت انتخاب شوند، با در نظر گرفتن تفکیکپذیریهای مکانی، طیفی و زمانی. علاوه بر این، دادههای پوشش گیاهی پیشبینیشده برای سناریوهای آینده که مدلهای EN را توانایی تولید نقشههای پیشبینی دقیقتر را ارائه میدهند، به آسانی در دسترس نیستند. این منجر به فرض مدل می شود، که ممکن است شرایط پوشش گیاهی مشابهی را هم در سناریوهای فعلی و هم در آینده داشته باشد که ممکن است قابلیت اطمینان مدل های پیش بینی شده را در پیش بینی ها کاهش دهد. علاوه بر این، اطلاعات مربوط به شرایط گلدهی که بر جمعیت زنبورها و آفات تأثیر می گذارد نیز به راحتی در دسترس نیست. نقشههای گل در کنیا تنها در گسترههای فضایی کوچکی موجود است که کل منطقه تحت این تحقیق را در بر نمیگیرد [ 65 ، 66]. به طور مشابه، حتی اگر زنبورستانها از سایتهای اکولوژیکی کشاورزی در کنیا انتخاب شدهاند، ممکن است سولههای آب و هوایی کامل آفات زنبور عسل مورد مطالعه قرار نگرفته باشند، به ویژه نقاط پسزمینه از کل وسعت کنیا ترسیم شدهاند. بنابراین، سهم متغیر فردی در گسترش آفات زنبور عسل در خارج از محدوده آموزشی آنها باید بیشتر مورد بررسی قرار گیرد.
5. نتیجه گیری ها
مدلهای EN توسعهیافته «خوب» بودند، از این رو برای پیشبینی مناسب بودن زیستگاه آفات زنبور عسل مناسب تلقی میشوند. با این حال، مشخص شد که مدل 1 که فقط از دادههای زیستاقلیمی استفاده میکند، مقادیر AUC کمتری با انحرافات استاندارد بالاتر نسبت به مدل 3 دارد که از دادههای زیستاقلیمی و سنجش از دور استفاده میکند. اگرچه دادههای سنجش از دور دقت مدلها را بهبود بخشید، متغیرهای زیست اقلیم بر مدلها بیش از متغیرهای سنجش از دور تأثیر گذاشتند. همچنین از دادههای زیستاقلیمی مدلسازیشده مشخص شد که تغییرات زیستاقلیمی بر توزیع این آفات زنبور عسل تأثیر خواهد داشت. توزیع فضایی آنها به مناطقی که در حال حاضر به عنوان کم وقوع طبقه بندی شده اند افزایش می یابد. با این حال، پیشبینی آفات پیشبینیشده میتواند بهبود یابد، اگر اقدامات مدیریت آفات مناسب انجام شود. از نقشه های پیش بینی می توان برای شناسایی مناطق پرخطر برای قرنطینه برای محدود کردن گسترش آفات استفاده کرد. بنابراین، این مدلها ابزار مهمی برای تصمیمگیرندگان در کاهش اثرات آفات مخرب خواهند بود. علاوه بر این، مناطق مهار نیاز به بررسی بیشتر برای ایجاد شرایطی دارند که تکثیر این آفات را منع کند. مداخلات هدفمند باید بر مناطقی متمرکز شود که در آنها شیوع آفات در آینده مورد انتظار است.
منابع
- کیاتوکو، ن. راینا، SK; مولی، ای. Mueke, J. افزایش کیفیت میوه در فلفل دلمه ای سالانه از طریق گرده افشانی توسط Hypotrigona gribodoi در Kakamega، کنیا غربی. انتومول. علمی 2014 ، 17 ، 106-110. [ Google Scholar ] [ CrossRef ]
- کلاین، A.-M. Vaissiere, BE; Cane, JH; استفان دیونتر، آی. کانینگهام، SA; کرمن، سی. Tscharntke, T. اهمیت گرده افشان ها در تغییر مناظر برای محصولات جهان. Proc. R. Soc. لندن بی: بیول. علمی 2007 ، 274 ، 303-313. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- راینا، SK; کیوکو، ای. زتنر، او. Wren، S. حفاظت از زیستگاه جنگل در آفریقا با استفاده از حشرات مهم تجاری. ان کشیش انتومول. 2011 ، 56 ، 465-485. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- مولی، ای. پچ، اچ. فریزر، ام. فریزر، جی. تورتو، بی. باومگارتن، تی. کیلونزو، جی. کیمانی، ج.ن. موموکی، اف. Masiga، D. ارزیابی توزیع و اثرات انگلها، پاتوژنها و آفتکشها بر روی جمعیت زنبور عسل ( Apis mellifera ) در شرق آفریقا. PLoS ONE 2014 , 16 , e94459. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- زاید، الف. ژنتیک زنبور عسل و حفاظت. Apidologie 2009 ، 40 ، 237-262. [ Google Scholar ] [ CrossRef ]
- Pirk، CWW; اشتراوس، یو. یوسف، ع.ا. دمارس، اف. انسان، H. سلامت زنبور عسل در آفریقا – بررسی. Apidologie 2015 ، 30 ، 1-25. [ Google Scholar ] [ CrossRef ]
- فومبونگ، AT; موموکی، FN; مولی، ای. Masiga، DK; آربوگاست، RT؛ تیل، PE; تورتو، ب. وقوع، تنوع و الگوی آسیب گونه Oplostomus (Coleoptera: Scarabaeidae)، آفات زنبور عسل در کنیا. Apidologie 2013 ، 44 ، 11-20. [ Google Scholar ] [ CrossRef ]
- تورتو، بی. فومبونگ، AT; Mutyambai، DM; مولی، ای. آربوگاست، RT؛ Teal، PEA Aethina tumida (Coleoptera: Nitidulidae) و Oplostomus haroldi (Coleoptera: Scarabaeidae): وقوع در کنیا، توزیع در کلنی های زنبور عسل، و پاسخ به بوهای میزبان. ان انتومول. Soc. صبح. 2010 ، 103 ، 389-396. [ Google Scholar ] [ CrossRef ]
- موموکی، FN; فومبونگ، آ. مولی، ای. Muigai، AWT؛ ماسیگا، دی. فهرستی از بیماری های مستند زنبورهای عسل آفریقایی. افر. انتومول. 2014 ، 22 ، 473-487. [ Google Scholar ] [ CrossRef ]
- پترسون، AT; Nakazawa، Y. مجموعه دادههای زیستمحیطی در مدلسازی طاقچههای زیستمحیطی مهم هستند: نمونهای با Solenopsis invicta و Solenopsis richteri . گلوب. Ecol. Biogeogr. 2008 ، 17 ، 135-144. [ Google Scholar ] [ CrossRef ]
- نویمان، پی. الیس، جی دی سوسک کندوی کوچک ( Aethina tumida Murray، Coleoptera: Nitidulidae): توزیع، زیست شناسی و کنترل یک گونه مهاجم. J. Apicult. Res. 2008 ، 47 ، 181-183. [ Google Scholar ] [ CrossRef ]
- نویمان، پی. پیرک، سی. هپبورن، اچ. سولبریگ، ا. راتنیکس، اف. الزن، پ. باکستر، جی. کپسولهسازی اجتماعی انگلهای سوسک توسط کلنیهای زنبور عسل کیپ ( Apis mellifera capensis Esch). Naturwissenschaften 2001 ، 88 ، 214-216. [ Google Scholar ] [ PubMed ]
- Simone-Finstrom، MD. Spivak, M. افزایش جمع آوری رزین پس از چالش انگل: موردی از خوددرمانی در زنبورهای عسل؟ PLoS ONE 2012 ، 7 ، e34601. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- پائو، اس. Gillespie، TW; Wolkovich، EM تشریح روابط غنای گونه های NDVI در جنگل های خشک هاوایی. J. Biogeogr. 2012 ، 39 ، 1678-1686. [ Google Scholar ] [ CrossRef ]
- فرناندز، م. همیلتون، اچ. قابلیت انتقال طاقچه اکولوژیکی با استفاده از گونه های مهاجم به عنوان مطالعه موردی. PLoS ONE 2015 ، 10 ، e0119891. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- کرنی، ام. پورتر، W. مدلسازی مکانیکی: ترکیب دادههای فیزیولوژیکی و فضایی برای پیشبینی محدوده گونهها. Ecol. Lett. 2009 ، 12 ، 334-350. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Champetier، A.; سامنر، دی. ویلن، جی اقتصاد زیستی زنبورهای عسل و گرده افشانی. محیط زیست منبع. اقتصاد 2014 ، 60 ، 143-164. [ Google Scholar ] [ CrossRef ]
- پترسون، AT; توپ، ال جی; Cohoon، KP پیش بینی توزیع پرندگان مکزیکی با استفاده از روش های مدل سازی طاقچه اکولوژیکی. بین المللی J. Avian Sci. 2002 ، 144 ، E27–E32. [ Google Scholar ] [ CrossRef ]
- فیلیپس، اس جی. Dudík، M. مدل سازی توزیع گونه ها با Maxent: الحاقات جدید و ارزیابی جامع. اکوگرافی 2008 ، 31 ، 161-175. [ Google Scholar ] [ CrossRef ]
- سیم، AF؛ کلاین، دی. Gernandt، DS; د لا روزا، JAP; دادههای سنجش از دور Dech، S. میتوانند پیشبینی غنای گونهها را با مدلهای توزیع گونهای انباشته بهبود بخشند: مطالعه موردی برای کاجهای مکزیکی. J. Biogeogr. 2014 ، 41 ، 736-748. [ Google Scholar ] [ CrossRef ]
- نویمان، پی. پتیس، جی اس. Schäfer, MO Quo vadis Aethina tumida ? بیولوژی و کنترل سوسک های کندو کوچک. Apidologie 2016 ، 47 ، 427-466. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- Speranza، CI; کیتم، بی. آمبنج، پ. ویزمن، یو. مکالی، اس. دانش بومی مربوط به تغییرپذیری و تغییر آب و هوا: بینش از خشکسالی در مناطق نیمه خشک ناحیه ماکونی سابق، کنیا. تغییر آب و هوا 2009 ، 100 ، 295-315. [ Google Scholar ] [ CrossRef ]
- گیتویی، اف. گیتاو، دبلیو. موتوآ، اف. Bauwens، W. تاثیر تغییر آب و هوا بر جریان شبیه سازی شده SWAT در غرب کنیا. بین المللی جی.کلیماتول. 2009 ، 29 ، 1823-1834. [ Google Scholar ] [ CrossRef ]
- دیتمن، وی. نازی، اف. مارتین، اس جی. اندرسون، دی.ال. لاک، بی. روش های استاندارد دلاپلان، KS برای تحقیقات واروآ. جی. آپیک. Res. 2013 ، 52 ، 1-54. [ Google Scholar ] [ CrossRef ]
- حردشت ع. باریوس، ام. فریفته، ج. میس، پی. کلمنت، جی. Verstraeten، WW; ترساگو، ک. ون رانست، م. کاپین، پی. برکمنز، دی. مدلسازی جایگاه اکولوژیکی بانک وولز در اروپای غربی. بین المللی جی. محیط زیست. Res. بهداشت عمومی 2013 ، 10 ، 499-514. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- مرو، سی. اسمیت، ام جی. Silander، JA راهنمای عملی MaxEnt برای مدلسازی توزیع گونهها: چه کاری انجام میدهد و چرا ورودیها و تنظیمات اهمیت دارند. اکوگرافی 2013 ، 36 ، 1058-1069. [ Google Scholar ] [ CrossRef ]
- اکلوندا، ال. Jönsson, P. TIMESAT 3.1 Manual Software ; دانشگاه لوند: لوند، سوئد، 2012. [ Google Scholar ]
- جانسون، پی. Eklundh, L. TIMESAT – برنامه ای برای تجزیه و تحلیل سری های زمانی داده های سنسور ماهواره ای. محاسبه کنید. Geosci. 2004 ، 30 ، 833-845. [ Google Scholar ] [ CrossRef ]
- کلارک، ام ال. دستیار، TM; Grau، HR; راینر، جی. یک رویکرد مقیاسپذیر برای نقشهبرداری پوشش سالانه زمین در 250 متر با استفاده از دادههای سری زمانی MODIS: مطالعه موردی در منطقه بومسازی چاکو خشک آمریکای جنوبی. سنسور از راه دور محیط. 2010 ، 114 ، 2816-2832. [ Google Scholar ] [ CrossRef ]
- جمالی، س. جانسون، پی. اکلوند، ال. آردو، جی. Seaquist، J. تشخیص تغییرات در روند پوشش گیاهی با استفاده از تقسیم بندی سری های زمانی. سنسور از راه دور محیط. 2015 ، 156 ، 182-195. [ Google Scholar ] [ CrossRef ]
- فوی، ا. تریمه، م. کاتکوونیک، وی. Egiazarian، K. مدل سازی و برازش نویز پواسونی-گاوسی عملی برای داده های خام تک تصویری. IEEE Trans. فرآیند تصویر 2008 ، 17 ، 1737-1754. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- شن، ی. وو، ال. دی، ال. یو، جی. تانگ، اچ. یو، جی. مدل های Shao, Y. Hidden Markov برای تخمین لحظه ای مراحل پیشرفت ذرت با استفاده از MODIS و داده های هواشناسی. Remote Sens. 2013 ، 5 ، 1734-1753. [ Google Scholar ] [ CrossRef ]
- فو، ایکس. Wang, L. کاهش ابعاد داده ها با کاربرد برای ساده سازی ساختار شبکه RBF و بهبود عملکرد طبقه بندی. IEEE Trans. سیستم مرد. سایبرن. بخش ب: سایبرن. 2003 ، 33 ، 399-409. [ Google Scholar ]
- هینتون، جنرال الکتریک؛ سالخوتدینوف، RR کاهش ابعاد داده ها با شبکه های عصبی. Science 2006 , 313 , 504-507. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- جارویس، ا. رویتر، HI; نلسون، ا. گوارا، E. SRTM پر از سوراخ برای جهان نسخه 4. پایگاه داده 90 متری CGIAR-CSI SRTM. 2008. در دسترس آنلاین: http://srtm.csi.cgiar.org/ (در 2 مارس 2015 قابل دسترسی است).
- Platts، PJ; Omeny، PA; Marchant، R. AFRICLIM: پیش بینی های آب و هوایی با وضوح بالا برای کاربردهای زیست محیطی در آفریقا. افر. جی. اکول. 2015 ، 53 ، 103-108. [ Google Scholar ] [ CrossRef ]
- Lovett, JC مدلسازی اثرات تغییر آب و هوا در آفریقا. افر. جی. اکول. 2015 ، 53 ، 1-2. [ Google Scholar ] [ CrossRef ]
- موالوسپو، اس. Tonnang، HEZ؛ ماساوه، ES؛ اوکوکو، برو؛ خدیولی، ن. یوهانسون، تی. کالاتایود، ص. Ru، BPL پیش بینی تاثیر تغییر دما بر توزیع آتی ساقه خواران ذرت و دشمنان طبیعی آنها در امتداد شیب کوهستان شرق آفریقا با استفاده از مدل های فنولوژی. PLoS ONE 2015 ، 10 ، e0130427. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- IPCC تغییرات آب و هوا 2013 – مبنای علوم فیزیکی: گروه کاری I در پنجمین گزارش ارزیابی پانل بین دولتی در مورد تغییرات آب و هوا. در هیئت بین دولتی تغییرات آب و هوایی ؛ انتشارات دانشگاه کمبریج: کمبریج، انگلستان، 2013; در دسترس آنلاین: http://ebooks.cambridge.org/ref/id/CBO9781107415324 (دسترسی در 3 اکتبر 2015).
- فیلیپس، اس جی. اندرسون، آر.پی. Schapire، RE مدلسازی حداکثر آنتروپی توزیعهای جغرافیایی گونهها. Ecol. مدل. 2006 ، 190 ، 231-259. [ Google Scholar ] [ CrossRef ]
- رودریگز-کاستاندا، جی. Hof، AR; جانسون، آر. هاردینگ، LE پیش بینی سرنوشت تنوع زیستی با استفاده از مدل های توزیع گونه ها: افزایش قابلیت مقایسه و تکرار مدل. PLoS ONE 2012 ، 7 ، e44402. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- پترسون، AT; پاپش، م. Eaton، M. قابلیت انتقال و ارزیابی مدل در مدلسازی طاقچه اکولوژیکی: مقایسه GARP و Maxent. اکوگرافی 2007 ، 30 ، 550-560. [ Google Scholar ] [ CrossRef ]
- وارد، جی. هستی، تی. بری، اس. الیت، جی. Leathwick، JR فقط داده های حضوری و الگوریتم EM. بیومتریک 2009 ، 65 ، 554-563. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Dormann، CF; الیت، جی. باچر، اس. بوخمن، سی. کارل، جی. کاره، جی. گارسیا مارکز، جی آر. گروبر، بی. لافورکید، بی. لیتائو، پی جی. و همکاران هم خطی: مروری بر روش های مقابله با آن و یک مطالعه شبیه سازی که عملکرد آنها را ارزیابی می کند. اکوگرافی 2013 ، 36 ، 27-46. [ Google Scholar ] [ CrossRef ]
- پترسون، AT; Cohoon، KP حساسیت الگوریتم های پیش بینی توزیعی به کامل بودن داده های جغرافیایی. Ecol. مدل. 1999 ، 117 ، 159-164. [ Google Scholar ] [ CrossRef ]
- Yackulic، CB; چندلر، آر. Zipkin، EF; رویل، جی. نیکولز، جی دی. کمپبل گرانت، EH; Veran, S. مدلسازی فقط حضوری با استفاده از MAXENT: چه زمانی میتوانیم به استنتاجها اعتماد کنیم؟ روش ها Ecol. تکامل. 2013 ، 4 ، 236-243. [ Google Scholar ] [ CrossRef ]
- دو، ز. وانگ، ز. لیو، ی. وانگ، اچ. ژو، اف. لیو، ی. مدلسازی طاقچه اکولوژیکی برای پیشبینی مناطق خطر بالقوه تب شدید با سندرم ترومبوسیتوپنی. بین المللی ج. عفونی کردن. دیس 2014 ، 26 ، 1-8. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Swets, JA اندازه گیری دقت سیستم های تشخیصی. علوم 1988 ، 240 ، 1285-1293. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Raes, N.; ter Steege، H. یک مدل تهی برای آزمایش اهمیت مدلهای توزیع گونههای فقط حضوری. اکوگرافی 2007 ، 30 ، 727-736. [ Google Scholar ] [ CrossRef ]
- هرناندز، PA; گراهام، CH; استاد، LL; آلبرت، DL اثر اندازه نمونه و ویژگیهای گونه بر عملکرد روشهای مدلسازی توزیع گونههای مختلف. اکوگرافی 2006 ، 29 ، 773-785. [ Google Scholar ] [ CrossRef ]
- مارتین، اس جی. هایفیلد، AC؛ برتل، ال. Villalobos، EM; بودجه، جنرال الکتریک؛ پاول، ام. شرودر، دی سی چشم انداز ویروسی زنبور عسل جهانی توسط یک کنه انگلی تغییر یافته است. علوم 2012 ، 336 ، 1304-1306. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- اشتراوس، یو. انسان، اچ. گوتیه، ال. کرو، آر.ام. دیتمن، وی. Pirk، CW شیوع فصلی پاتوژن ها و انگل ها در زنبور عسل ساوانا ( Apis mellifera scutellata ). J. Invertebr. پاتول. 2013 ، 114 ، 45-52. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- راینا، SK; Kimbu، DM تغییرات در نژادهای زنبور عسل Apis mellifera (Hymenoptera: Apidae) در کنیا. بین المللی جی تروپ. Insect Sci. 2005 ، 25 ، 281-291. [ Google Scholar ] [ CrossRef ]
- وایلی، EO; McNyset، KM; پترسون، AT; رابینز، CR; دیدگاه مدلسازی استوارت، AM طاقچه در مورد پیش بینی محدوده جغرافیایی در محیط دریایی با استفاده از یک الگوریتم یادگیری ماشینی. اقیانوس شناسی 2003 ، 16 ، 120-127. [ Google Scholar ] [ CrossRef ]
- مانی، MS اکولوژی و جغرافیای زیستی حشرات ارتفاعات ; Springer Science & Business Media: برلین، آلمان، 2013. پ. 539. [ Google Scholar ]
- آراندا، SC; Lobo, JM مدلسازی توزیع گونههای مبتنی بر حضور چقدر به خوبی تنوع مجموعه را پیشبینی میکند؟ مطالعه موردی فلور تنریف. اکوگرافی 2011 ، 34 ، 31-38. [ Google Scholar ] [ CrossRef ]
- ساعتچی، س. بوئرمن، دبلیو. تر استیج، اچ. موری، س. اسمیت، سل مدلسازی توزیع گونه های درختی آمازون و تنوع با استفاده از اندازه گیری های سنجش از دور. سنسور از راه دور محیط. 2008 ، 112 ، 2000–2017. [ Google Scholar ] [ CrossRef ]
- آتور، اف. آلفو، م. دی سانکتیس، ام. فرانچسکونی، اف. Bruno, F. مقایسه روشهای درونیابی برای نقشه برداری متغیرهای اقلیمی و زیست اقلیمی در مقیاس منطقه ای. بین المللی جی.کلیماتول. 2007 ، 27 ، 1825-1843. [ Google Scholar ] [ CrossRef ]
- Hijmans، RJ; کامرون، SE; پارا، جی ال. جونز، پی.جی. جارویس، A. سطوح آب و هوایی درونیابی شده با وضوح بسیار بالا برای مناطق خشکی جهانی. بین المللی جی.کلیماتول. 2005 ، 25 ، 1965-1978. [ Google Scholar ] [ CrossRef ]
- انگلر، آر. گیسان، ع. Rechsteiner, L. یک رویکرد بهبود یافته برای پیشبینی توزیع گونههای کمیاب و در خطر انقراض از دادههای وقوع L و شبه عدم وجود. J. Appl. Ecol. 2004 ، 41 ، 263-274. [ Google Scholar ] [ CrossRef ]
- بردلی، کارشناسی; اولسون، AD; وانگ، او. دیکسون، بی جی; پلک، ال. Sesnie، SE; Zachmann، LJ تشخیص گونه در مقابل تناسب زیستگاه: آیا ما مدلهای مناسب زیستگاه را با دادههای سنجش از دور سوگیری میکنیم؟ Ecol. مدل. 2012 ، 244 ، 57-64. [ Google Scholar ] [ CrossRef ]
- کلیفورد، جی دی. Tarassenko, L. کمی سازی خطاها در تخمین های طیفی HRV به دلیل جایگزینی ضربان و نمونه برداری مجدد. IEEE Trans. بیومد. مهندس 2005 ، 52 ، 630-638. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- دیکشیت، او. روی، DP بررسی تجربی اثرات نمونهبرداری مجدد تصویر بر طبقهبندی نظارت شده طیفی و بافتی یک تصویر چند طیفی با وضوح فضایی بالا. فتوگرام مهندس Remote Sens. 1996 , 62 , 1085-1092. [ Google Scholar ]
- فیلدینگ، ق. Bell, JF مروری بر روشهای ارزیابی خطاهای پیشبینی در مدلهای حضور/غیاب حفاظتی. محیط زیست حفظ کنید. 1997 ، 24 ، 38-49. [ Google Scholar ] [ CrossRef ]
- عبدالرحمن، EM; ماکوری، دی.م. لندمن، تی. پیروینن، آر. قاسم، س. پلیککا، پ. Raina, SK کاربرد داده های فراطیفی عقاب AISA و طبقه بندی تصادفی جنگل برای نقشه برداری گل. Remote Sens. 2015 ، 7 ، 13298–13318. [ Google Scholar ] [ CrossRef ]
- لندمن، تی. پیروینن، آر. ماکوری، دی.م. عبدالرحمن، EM; ماکائو، اس. پلیککا، پ. Raina, SK کاربرد سنجش از دور فراطیفی برای نقشه برداری گل در ساوانای آفریقا. سنسور از راه دور محیط. 2015 ، 166 ، 50-60. [ Google Scholar ] [ CrossRef ]
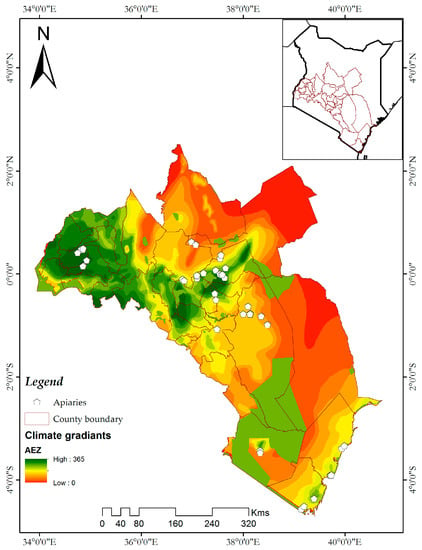
شکل 1. منطقه مورد مطالعه و توزیع زنبورستان ها (نقاط سفید) که از آنجا داده های وقوع در چهار منطقه در سراسر کنیا طبقه بندی شده توسط شیب آب و هوای نماینده جمع آوری شد. پس زمینه شیب های آب و هوایی در سراسر منطقه را نشان می دهد. زنبورستان ها در مناطق مختلف کشاورزی توزیع شده اند.

شکل 2. ماتریس همخطی برای متغیرهای پیش بینی کننده مدل های طاقچه اکولوژیکی. آستانه هم خطی در |r| تعیین شد > 0.7 بر اساس Dormann و همکاران. [ 44 ]. سایههای تیرهتر رنگهای آبی و قرمز نشاندهنده همخطی متغیر زیاد است در حالی که سایههای روشنتر نشاندهنده همخطی بودن کم است. متغیرهای زیست اقلیمی، به ویژه bio12 (میانگین بارندگی سالانه) و dm (تعداد ماههای خشک) نسبت به متغیرهای زیستی و توپوگرافی همخطی بالایی از خود نشان دادند.

شکل 3. خلاصه گرافیکی روش شناسی که تعامل بین متغیرهای مختلف مورد استفاده برای ساخت سه مدل آفت زنبور عسل و انتخاب مدل مناسب را نشان می دهد.
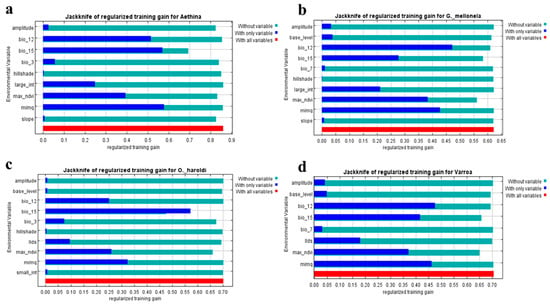
شکل 4. سهم متغیر Jackknife برای ( a ) A. tumida ، ( b ) G. mellonella ، ( c ) O. haroldi و ( d ) V. destructor برای سنجش از دور و مدل زیست اقلیم (مدل 3). سایه های آبی تیره افزایش تمرین منظم شده را برای متغیر خاص، آبی روشن بدون متغیر در حالی که قرمز افزایش تمرین منظم را با همه متغیرها نشان می دهد.
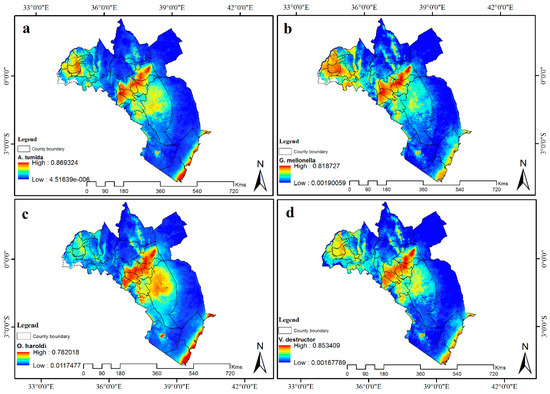
شکل 5. تناسب زیستگاه فعلی برای ( الف ) A. tumida ، ( ب ) G. mellonella ، ( c ) O. haroldi و ( d ) V. destructor در کنیا. آبی نشان دهنده تناسب زیستگاه کم است در حالی که رنگ قرمز نشان دهنده تناسب زیستگاه بالا است. کوه کنیا و مناطق ساحلی برای همه آفات در معرض خطر بالا پیش بینی شده بود در حالی که خطر در Kakamega از یک آفت به آفت دیگر متفاوت بود.
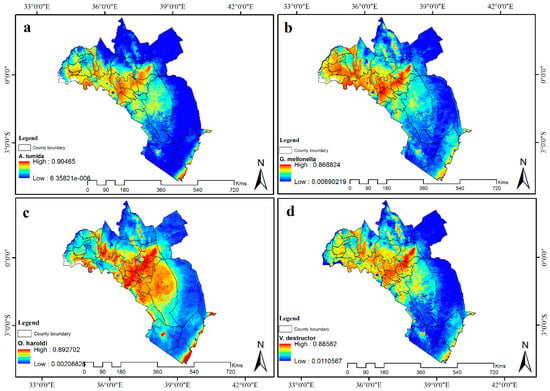
شکل 6. مناسب بودن زیستگاه پیش بینی شده (2055) برای ( a ) A. tumida ، ( b ) G. mellonela ، ( c ) O. haroldi و ( d ) V. destructor در کنیا. آبی نشان دهنده تناسب زیستگاه کم است در حالی که رنگ قرمز نشان دهنده تناسب زیستگاه بالا است. پیشبینی میشود مناطق پرخطر از نظر فضایی در سراسر کشور برای همه آفات زنبور عسل افزایش یابد.
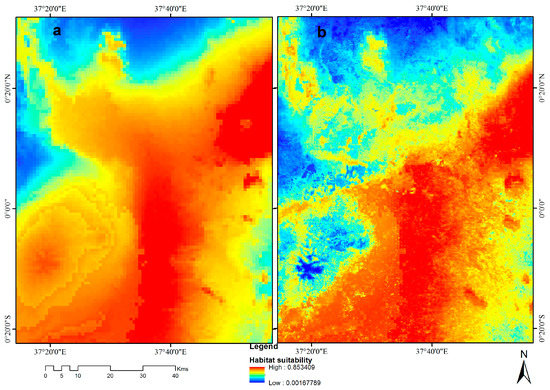
شکل 7. بهبود پیش بینی و کاهش بیش از حد برازش مدل های اکولوژیکی با استفاده از متغیرهای سنجش از دور. نقشه سمت چپ ( a ) زیرمجموعه ای از مدل بیوکلیماتیک V. destructor (مدل 1) است که تخمین بیش از حد این آفت زنبور عسل را در قسمت وسط و سمت چپ نقشه در مقایسه با مدل سنجش از دور و زیست اقلیم (مدل 3) نشان می دهد. ( ب ) که پیشبینی بالایی را نشان میدهند که به گوشه سمت راست پایین نقشه کاهش یافته است.
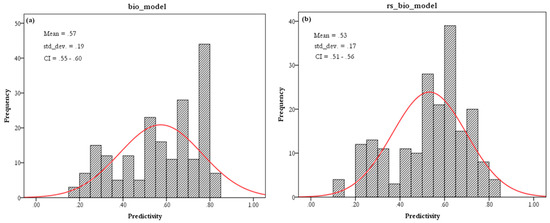
شکل 8. فرکانس پیش بینی ( الف ) مدل های بیولوژیکی (مدل 1) و ( ب ) سنجش از دور و مدل های بیولوژیکی (مدل 3) برای V. destructor . هیستوگرام سنجش از دور و مدل زیست اقلیم (مدل 3) دارای میانگین و انحراف معیار (SD) کمتری نسبت به مدل زیست اقلیم (مدل 1) بود. پیش بینی برای مدل سنجش از دور و زیست اقلیم (مدل 3) حول میانگین متمرکز شد در حالی که پیش بینی مدل های زیست اقلیم (مدل 1) به سمت راست منحرف شد.

جدول 1. متغیرهای مورد استفاده در مدل طاقچه اکولوژیکی به متغیرهای زیستی سنجش از راه دور که از دادههای شاخص گیاهی تفاوت نرمال شده در فضا (NDVI) مشتق شدهاند، خوشهبندی شدند. متغیرهای توپوگرافی از مدل رقومی ارتفاع از ماموریت توپوگرافی رادار شاتل (SRTM) و متغیرهای زیست اقلیم از Africlim استخراج شدند.
جدول 2. میانگین فراوانی آفت ثبت شده در طول دوره نظارت در فصل مرطوب و خشک در کنیا. بر اساس آزمون من ویتنی، تعداد میانگین ها در هر ستون با حرف مشابه تفاوت معنی داری (05/0 ≤ p) با یکدیگر ندارند.
جدول 3. مساحت زیر منحنی (AUC) و انحراف استاندارد (SD) مدلهای بیوکلیماتیک (مدل 1)، سنجش از دور (مدل 2) و سنجش از دور و مدلهای زیستاقلیمی (مدل 3) برای A. tumida ، G. mellonella ، O. هارولدی و وی ویرانگر .
جدول 4. سهم، به عنوان درصد، متغیرهای مختلف زیست اقلیمی و سنجش از دور در چهار مدل طاقچه اکولوژیکی با استفاده از jackknife. سهم کل خوشه در پایین خوشه ها برای هر گونه آفت نشان داده شده است.
© 2017 توسط نویسندگان. دارنده مجوز MDPI، بازل، سوئیس. این مقاله یک مقاله با دسترسی آزاد است که تحت شرایط و ضوابط مجوز Creative Commons Attribution (CC BY) ( http://creativecommons.org/licenses/by/4.0/ ) توزیع شده است.
بدون نظر