

چکیده
فعالیتهای نظارت زیستی بر کسب و تجزیه و تحلیل دادههای اپیدمیولوژیک و بیولوژیکی برای تفسیر رویدادهای آشکار و پیشبینی نتایج در شیوع بیماریهای عفونی متمرکز است. ما یک چارچوب مدلسازی ریاضی را بر اساس منابع دادهای همسو با جغرافیایی و با انعطافپذیری مناسب توصیف میکنیم که مدلسازی بیماری را به دو سطح مجزا اما همراه تقسیم میکند. یک شبیهسازی تصادفی سطح بالا در شبکهای با گرههایی تعریف میشود که «وصلههای» جغرافیایی قابل تنظیم توسط کاربر را نشان میدهند. گسترش بیماری درون پچ با معادلات دیفرانسیل که اختلاط یکنواخت درون پچ را فرض می کنند درمان می شود. ما از داده های جمع آوری شده در سطح شهرستان ایالات متحده در مورد جمعیت حیوانات و پارامترهای موجود در ادبیات برای شبیه سازی گسترش اپیدمی دو عامل بیماری های حیوانی متفاوت استفاده می کنیم: بیماری تب برفکی و آنفولانزای فوق حاد پرندگان. نتایج نشاندهنده توانایی این چارچوب برای استفاده از دادههای با وفاداری پایین در حین تولید خروجی معنیدار برای اطلاع از نظارت زیستی و اقدامات کنترل بیماری است. به عنوان مثال، ما نشان میدهیم که بزرگی احتمالی یک شیوع نسبت به محل شروع شیوع حساس است و وابستگی شدید جغرافیایی اپیدمیهای بیماریهای عفونی دام و طیور و سودمندی سیاست نظارت زیستی مؤثر را برجسته میکند. توانایی مقایسه بیماریهای مختلف و جمعیت میزبان در سراسر چشمانداز جغرافیایی برای برنامههای پشتیبانی تصمیم و برای ارزیابی تأثیر پروتکلهای نظارت، تشخیص و کاهش اهمیت دارد. نتایج نشاندهنده توانایی این چارچوب برای استفاده از دادههای با وفاداری پایین در حین تولید خروجی معنیدار برای اطلاع از نظارت زیستی و اقدامات کنترل بیماری است. به عنوان مثال، ما نشان میدهیم که بزرگی احتمالی یک شیوع نسبت به محل شروع شیوع حساس است و وابستگی شدید جغرافیایی اپیدمیهای بیماریهای عفونی دام و طیور و سودمندی سیاست نظارت زیستی مؤثر را برجسته میکند. توانایی مقایسه بیماریهای مختلف و جمعیت میزبان در سراسر چشمانداز جغرافیایی برای برنامههای پشتیبانی تصمیم و برای ارزیابی تأثیر پروتکلهای نظارت، تشخیص و کاهش اهمیت دارد. نتایج نشاندهنده توانایی این چارچوب برای استفاده از دادههای با وفاداری پایین در حین تولید خروجی معنیدار برای اطلاع از نظارت زیستی و اقدامات کنترل بیماری است. به عنوان مثال، ما نشان میدهیم که بزرگی احتمالی یک شیوع نسبت به محل شروع شیوع حساس است و وابستگی شدید جغرافیایی اپیدمیهای بیماریهای عفونی دام و طیور و سودمندی سیاست نظارت زیستی مؤثر را برجسته میکند. توانایی مقایسه بیماریهای مختلف و جمعیت میزبان در سراسر چشمانداز جغرافیایی برای برنامههای پشتیبانی تصمیم و برای ارزیابی تأثیر پروتکلهای نظارت، تشخیص و کاهش اهمیت دارد. ما نشان میدهیم که بزرگی احتمالی یک شیوع نسبت به محل شروع شیوع حساس است و وابستگی شدید جغرافیایی اپیدمیهای بیماریهای عفونی دام و طیور و سودمندی سیاست نظارت زیستی مؤثر را برجسته میکند. توانایی مقایسه بیماریهای مختلف و جمعیت میزبان در سراسر چشمانداز جغرافیایی برای برنامههای پشتیبانی تصمیم و برای ارزیابی تأثیر پروتکلهای نظارت، تشخیص و کاهش اهمیت دارد. ما نشان میدهیم که بزرگی احتمالی یک شیوع نسبت به محل شروع شیوع حساس است و وابستگی شدید جغرافیایی اپیدمیهای بیماریهای عفونی دام و طیور و سودمندی سیاست نظارت زیستی مؤثر را برجسته میکند. توانایی مقایسه بیماریهای مختلف و جمعیت میزبان در سراسر چشمانداز جغرافیایی برای برنامههای پشتیبانی تصمیم و برای ارزیابی تأثیر پروتکلهای نظارت، تشخیص و کاهش اهمیت دارد.
کلید واژه ها:
اپیدمیولوژی فضایی ; بیماری تب برفکی ؛ آنفلوانزای مرغی H5N1 ؛ نظارت زیستی ; شبیه سازی اپیدمی ; جغرافیا
1. مقدمه
بیماریهای عفونی نوظهور از منظر اقتصاد جهانی، امنیت و سلامت عمومی یک نگرانی اساسی هستند. اکثر بیماری های عفونی جدید مهم برای سلامت عمومی از مخازن حیوانات پدید آمده اند و به عنوان مشترک بین انسان و دام در نظر گرفته می شوند [ 1 ]. سایر اپیدمی های حیوانات مانند شیوع بیماری تب برفکی (FMD) در سال 2001 در بریتانیا منجر به معدوم کردن تقریباً چهار میلیون حیوان و هزینه ای بین 3.7-6.3 میلیارد دلار شد [ 2 ]. گسترش اپیزوتیک کنونی آنفولانزای پرندگان بسیار بیماری زا (HPAI) یک زیرگروه H5N1 در میان گونه های پرندگان وحشی و طیور اهلی منجر به نابودی 200 میلیون پرنده با تأثیری بالغ بر 10 میلیارد دلار [ 3 ] شده است و همچنان به عنوان یک تهدید مشترک بین انسان و دام ادامه دارد [ 4 ].]. تا پایان سال 2012، 610 عفونت آنفلوانزای H5N1 انسانی در سراسر جهان با 59 درصد مرگ و میر وجود داشته است [ 5 ]. ظهور سویه آنفولانزای همه گیر A (H1N1) در سال 2009 به عنوان یک دسته بندی مجدد سه گانه از ویروس های آنفلوانزای خوکی بر نیاز به نظارت فشرده در جمعیت دام برای بیماری های عفونی آینده با پتانسیل مشترک بین انسان و دام تاکید می کند [ 6 ]. تشخیص زودهنگام تهدیدات بیماریهای عفونی اضطراری، که با تلاشهای نظارتی پیشرفته امکانپذیر میشود، میتواند عوارض پاییندستی، مرگومیر و پیامدهای اقتصادی ناشی از شیوع بیماریهای عفونی در مقیاس بزرگ را کاهش دهد.
فعالیتهای نظارت زیستی بر کسب و تجزیه و تحلیل دادههای اپیدمیولوژیک و بیولوژیکی برای تفسیر رویدادهای آشکار و پیشبینی نتایج در شیوع بیماریهای عفونی متمرکز است. در ساعات اولیه و روزهای یک رویداد بیولوژیکی جدید، دادههای مربوط به پارامترهای حیاتی مانند تعداد عفونتهای ثانویه ایجاد شده توسط یک مورد اولیه و زمان تولید سریال معمولاً خام هستند [ 7 ]. کیفیت داده ها به طور کلی با پیشرفت رویداد بهبود می یابد. عدم قطعیت بزرگ در تخمین تعداد پایه تولید مثل در روزهای اولیه اپیدمی سارس در سال 2003 در پکن با پیشرفت شیوع کاهش یافت [ 8 ]. به طور مشابه، نسبت مرگ و میر مربوط به سویه همه گیر آنفولانزای A H1N1 در سال 2009 در ابتدا با دامنه وسیعی از 0.8٪ تا 1.8٪ برآورد شد.9 ]. Garske و همکاران [ 10 ] تغییر در تشخیص موارد از موارد خفیف به موارد شدید را با پیشرفت همهگیری و تاخیر گزارش بین شروع علائم و مرگ در روزهای اولیه رویداد به عنوان دلایل اولیه برای ارزیابی شدت بیماری شناسایی کردند. بنابراین، مدل هایی که با اطلاعات محدود، می توانند راهنمایی های عملی و طیفی از نتایج ممکن را تحت سناریوهای کنترل و نظارت مختلف ارائه دهند، می توانند ابزار ارزشمندی برای دانشمندان و سیاست گذاران باشند.
پیشرفت در علم و فناوری اطلاعات باعث ایجاد قابلیتهای جمعآوری دادههای بهداشت عمومی در زمان واقعی میشود و نیاز به «مدلهای کمک تصمیمگیری» را الزامی میکند که میتوانند به دادههای نظارتی مرتبط شوند و در طول یک اپیدمی بازخورد بلادرنگ ارائه دهند [ 11 ]. این مدلها از سیاستگذاران پشتیبانی میکنند و تصمیمات آنها را در مورد زمان، مکان، و شدت اجرای اقدامات کنترلی مانند قرنطینه، محدودیتهای حرکتی، استراتژیهای واکسن، و معدومسازی در مورد بیماریهای حیوانات آگاه میکنند.
از آنجایی که همهگیریها میتوانند از چندین سیستم میزبان پاتوژن شناختهشده و ناشناخته ناشی شوند، مدلهایی که از فعالیتهای نظارت زیستی پشتیبانی میکنند باید ماهیت انعطافپذیر و پویا داشته باشند و بتوانند دادههای جدید و با کیفیت بهتر را با تغییر موقعیت ترکیب کنند. تغییرات در پارامترها، مانند جهش ویروس که منجر به بیماریزایی تغییر میشود، یا تغییر در توزیع میزبان یا صلاحیت، باید به راحتی در چارچوب مدل موفق گنجانده شود. در برخی موارد، ساختار مسیرهای انتقال ممکن است نیاز به تغییر داشته باشد تا ویژگیهای کیفی جدید شیوع در حال پیشرفت را تطبیق دهد. به عنوان مثال، یک مسیر انتقال ناشناخته قبلی برای انتقال پاتوژن ممکن است کشف شود و نیاز به گنجاندن داشته باشد. در شروع یک اپیدمی، پارامترها اغلب بسیار نامشخص هستند، اما عدم قطعیت را می توان با پیشرفت اپیدمی کاهش داد و داده های بهتری در دسترس قرار گرفت و درک دقیق تری از مکانیسم های علت شناختی پدیدار شد. یک چارچوب مدل ایده آل باید بتواند با تغییر وضعیت دانش تکامل یابد.
ما یک رویکرد مدلسازی ریاضی را توصیف میکنیم که به اندازه کافی انعطافپذیر است تا تنوع سیستمهای میزبان پاتوژن و وضعیت در حال تکامل دانش در مورد سیستمها را در خود جای دهد. انعطافپذیری با تقسیمبندی مدلسازی گسترش و کاهش بیماری به دو سطح مجزا اما همراه به دست میآید. اول یک شبیهسازی تصادفی سطح بالا با تکهها (یا گرهها) است که از طریق یک شبکه تماسی تعریفشده بر روی یک شبکه [ 12 ] متشکل از تمام فعل و انفعالات کوتاه و بلند مربوط به انتقال بیماری هستند. درون هر گره یک سطح دوم “مخفی” درون یک پچ وجود دارد که در آن گسترش بیماری از معادلات دیفرانسیل معمولی SIR مانند میدان متوسط پیروی می کند [ 13 ]]. این چارچوب استراتژیهای کاهش را در هر دو مدل سطح بالا و پایین ترکیب میکند و امکان سیاستهای متنوعی مانند کنترل حرکت، نظارت، قرنطینه، واکسن، و حذف را فراهم میکند. در نهایت، با توجه به سطوح متعدد عدم قطعیت در شیوع بیماری، ما تصادفی را در گسترش فضایی گنجاندهایم و مدل را چندین بار با تغییر در مقادیر پارامتر و مکان اولیه شیوع اجرا کردیم. این کمی سازی عدم قطعیت طیف قابل قبولی از نتایج را برای تصمیم گیرندگان فراهم می کند.
چارچوبی که ما در اینجا طرح میکنیم بر اساس کار موجود و گسترش آن برای مدلسازی گسترش فضایی بیماریهای حیوانی [ 14 ، 15 ، 16 ، 17 ، 18 ، 19 ، 20 ] است. کار ما نقشهای مهم و متقابل جغرافیا و جنبههای فضایی تنوع میزبان، توزیع میزبان، حرکت در فواصل دور و کوتاه، تعامل انسان و دام و مداخلات احتمالی را برجسته میکند.
چارچوب مدلی که در اینجا توضیح داده شده است، توانایی مدیریت طیف گستردهای از گونههای میزبان و محرکهای قابلتوجه شیوع بیماری ناشی از زیستشناسی میزبان-پاتوژن (مثلاً طول دوره بدون علائم بالینی)، اقدامات کنترلی (مانند تأثیر موضعی در مقابل. کنترلهای جهانی حرکت)، و وابستگی جغرافیایی (آغاز شیوع در مناطقی با تولید دام متراکم در مقابل محیطهای روستایی بیشتر) که ممکن است فرصتهایی را برای کاهش شیوع فراهم کند. نشان داده شده است که ترکیب ساختار فضایی شبکه تماس میزبان حساس برای پیشبینی پارامترهای اپیدمیولوژیک در مقیاس جمعیتی مانند تعداد اولیه تولید مثل و نرخ حمله مهم است [ 21 ، 22 ، 23 ]]. مدل های بسیار دقیق FMD 2001 در انگلستان توسط Keeling و همکاران. [ 23 ] و فرگوسن و همکاران. [ 14 ] به دلیل در دسترس بودن داده های موقعیت جغرافیایی دقیق در مزارع دام امکان پذیر بود. دادههای سرشماری کشاورزی در دسترس عموم در مورد جمعیت دام در ایالات متحده فقط به صورت انبوه در سطح شهرستان وجود دارد. مدلی که در اینجا توضیح داده میشود را میتوان در سطح شهرستان برای ارائه اطلاعات و بینش حتی با کمبود دادههای مکان مزرعه و حرکت دام با کیفیت بالا، مقیاسبندی کرد.
در این کار، کلیت و سودمندی این چارچوب را با ارائه دو برنامه کاربردی از مدل خود، شبیهسازی اپیدمی دو بیماری مهم اقتصادی حیوانی با میزبانهای مختلف، برجسته میکنیم: بیماری تب برفکی (FMD) و آنفولانزای پرندگان بسیار بیماریزا (HPAI) . FMD و HPAI در جمعیتهای میزبان حیوانات بسیار متفاوت (گاو، گوسفند و خوک برای FMD و طیور برای HPAI) گسترش مییابند و ویژگیهای علتشناسی دوگانه دارند (FMD: تاریخ طبیعی وابسته به گونه، مرگومیر کم، پتانسیل بالا برای بومی شدن و HPAI: همگن تاریخ طبیعی، نزدیک به 100٪ مرگ و میر). ما فقط از دادههای جمعیت دام در دسترس عموم استفاده میکنیم که در سطح شهرستان جمعآوری شده و هر پنج سال یک بار توسط سرویس ملی آمار کشاورزی بهروزرسانی میشود [ 24 ].
2. مواد و روشها
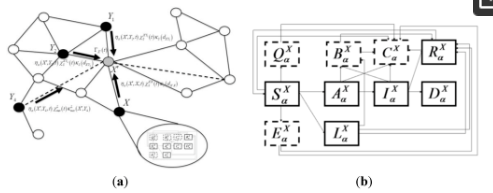
این مدل یک شبیهسازی مبتنی بر فضایی است که از نظر جغرافیایی در سطح شهرستان در ایالات متحده و شبیه به آنچه قبلاً توضیح داده شد [ 25 ] است. مدل سطح بالا («بین پچ») یک شبیهسازی تصادفی است که در آن هر «پچ» یک واحد جغرافیایی یا اپیدمیولوژیکی است که در آن اختلاط یکنواخت ( یعنی نرخهای تماس بین میزبان همگن) فرض میشود. شماتیکی که ساختار مدل را با حالت هایی که به صورت جعبه و انتقال ها به صورت فلش نشان داده شده است در شکل 1 نشان داده شده است . هر جمعیت میزبان حساس در یک لکه X و گونه (یا نوع دام) a از تاریخ طبیعی خاص بیماری که در شکل 1 نشان داده شده است، پیروی می کند .
ما با مدل بین پچ برای گسترش جغرافیایی شروع می کنیم. در زمان t در طول شبیه سازی، اگر یک پچ X خاص حساس باشد (به عنوان مثال ، بدون میزبان های آلوده)، احتمال آلوده شدن آن در زمان t توسط داده می شود.
p X ( t ) = 1 − exp(−Γ X ( t ))
هسته، Γ X ( t )، که گسترش عفونت را توصیف می کند، تابعی از فاصله از لکه های آلوده نزدیک و ارتباط با لکه های آلوده دور از طریق حرکت دام است. هسته عفونت شهرستان همچنین به اقدامات کاهشی مربوط به کاهش جابجایی دام در مقیاس های مسافت طولانی یا کوتاه وابسته است. حالت های “درون پچ” مدل بیماری، تاریخچه طبیعی بیماری را در میزبان گونه ها یا دام های نوع a در شهرستان (یا پچ) X مشخص می کند. بخش های بیماری شامل میزبان های حساس ( ) ، میزبان های آلوده بدون علامت که به علائم پیشرفت می کنند ( ) ، میزبان های آلوده بدون علامت که به علائم پیشرفت نمی کنند ( یعنی  .، یک حالت ناقل)، ( )، میزبان های آلوده علامت دار ( )، میزبان های بهبود یافته که عفونت را به طور طبیعی یا از طریق ایمن سازی پاک کرده اند ( ) و میزبان های آلوده ای که در اثر این بیماری مرده اند ( ). مانند هر مدل بیماری جزئی، بسته به سیستم بیماری در نظر گرفته شده می توان به راحتی بخش ها را اضافه یا کم کرد.
.، یک حالت ناقل)، ( )، میزبان های آلوده علامت دار ( )، میزبان های بهبود یافته که عفونت را به طور طبیعی یا از طریق ایمن سازی پاک کرده اند ( ) و میزبان های آلوده ای که در اثر این بیماری مرده اند ( ). مانند هر مدل بیماری جزئی، بسته به سیستم بیماری در نظر گرفته شده می توان به راحتی بخش ها را اضافه یا کم کرد.
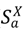
 .، یک حالت ناقل)، ( )، میزبان های آلوده علامت دار ( )، میزبان های بهبود یافته که عفونت را به طور طبیعی یا از طریق ایمن سازی پاک کرده اند ( ) و میزبان های آلوده ای که در اثر این بیماری مرده اند ( ). مانند هر مدل بیماری جزئی، بسته به سیستم بیماری در نظر گرفته شده می توان به راحتی بخش ها را اضافه یا کم کرد.
.، یک حالت ناقل)، ( )، میزبان های آلوده علامت دار ( )، میزبان های بهبود یافته که عفونت را به طور طبیعی یا از طریق ایمن سازی پاک کرده اند ( ) و میزبان های آلوده ای که در اثر این بیماری مرده اند ( ). مانند هر مدل بیماری جزئی، بسته به سیستم بیماری در نظر گرفته شده می توان به راحتی بخش ها را اضافه یا کم کرد.

حالت هایی نیز با معماری کاهش/پاسخ وجود دارد. علاوه بر کنترلهای حرکتی مورد بحث، قرنطینه میزبانهای حساس، واکسیناسیون میزبانهای حساس (پیشگیرانه) و آلوده (درمان برای کاهش عفونت) و کشتن میزبانهای حیوانی را در نظر میگیریم. ایالت های مربوط به اقدامات کنترلی، مستعدهایی هستند که با موفقیت قرنطینه شده اند ( )، افراد مستعدی که با موفقیت واکسینه شده اند و هنوز هم می توانند با نرخ کمتری برای مدت کوتاهی قبل از انتقال به کلاس بهبودیافته آلوده شوند ( ) میزبان های آلوده ای که با واکسیناسیون با موفقیت درمان شده اند و حیواناتی که معدوم شده اند. ). پویایی انتقال و پیشرفت بیماری توسط سیستم معادلات جفت شده به تفصیل در پیوست توضیح داده شده است.
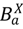
 ). پویایی انتقال و پیشرفت بیماری توسط سیستم معادلات جفت شده به تفصیل در پیوست توضیح داده شده است.
). پویایی انتقال و پیشرفت بیماری توسط سیستم معادلات جفت شده به تفصیل در پیوست توضیح داده شده است.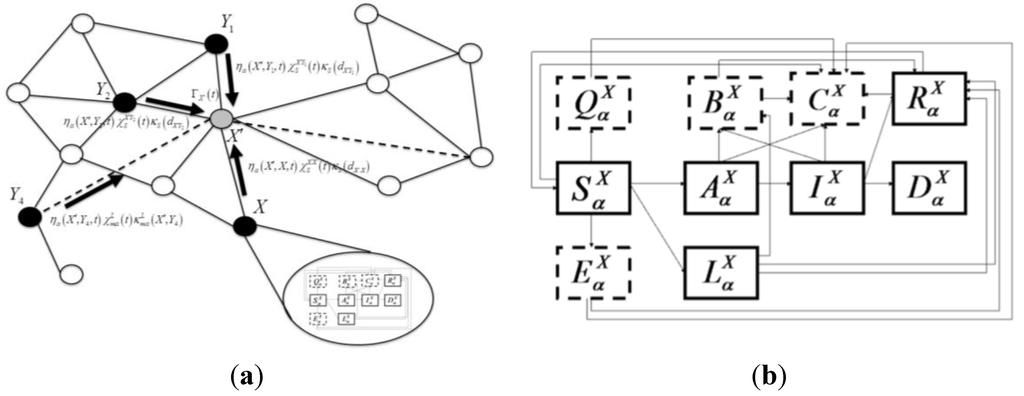
شکل 1. شماتیک برای مدل انتقال و پیشرفت بیماری در داخل شهرستان. جعبه ها وضعیت حیوانات گونه a در شهرستان X را نشان می دهند. جعبههای جامد وضعیتهای بیماری سابقه طبیعی را برچسبگذاری میکنند: حساس ( )، بدون علائم بالینی آلوده ( )، وضعیت ناقل پنهان آلوده ( )، عفونی علامتدار ( )، بهبود یافته ( ) و مرده ( ) (فقط به دلیل مرگ و میر بیماری). کشورهایی که با خطوط چین مرزی شده اند، حالت های کاهش دهنده را نشان می دهند: مستعد قرنطینه ( )، مستعد واکسینه شده به صورت پیشگیرانه ( )، حیوانات آلوده واکسینه شده ( ) و حیوانات معدوم شده ( ).
معماری اندازهگیری نظارت/کنترل در تاریخچه طبیعی بیماری توسط توابع «دروازهای» کنترل میشود که بسته به زمان مقادیر صفر یا یک را میگیرند و تعیین میکنند که چه زمانی کاهشدهنده روشن و خاموش میشود . اثربخشی کاهش به عنوان درجه موفقیتی که هنگام اجرا به دست می آید تعریف می شود. اینها توسط پارامترها داده می شود . به عنوان مثال، در معادله S3a (تکمیلی)، این اصطلاح سرعت قرنطینه شدن افراد مستعد را توصیف می کند. در حالی که نرخ انتقال تعداد میزبانهای حساسی را که احتمالاً میتوانند در واحد زمان قرنطینه شوند، تعیین میکند، پارامتر کیفیت اندازه گیری کنترل را تعیین می کند. پارامترهای اندازه گیری کنترل می تواند تابعی از نوع و مکان گونه باشد، بنابراین اقدامات کنترلی بسیار خاص را می توان ایجاد کرد. برای واکسیناسیون، ما فرض می کنیم که زیست شناسی اساسی عامل اثربخشی غالب است، بنابراین این متغیرها وابسته به مکان نیستند. ما فرض می کنیم که تمام حیوانات تعیین شده برای معدوم سازی معدوم خواهند شد، بنابراین اثربخشی را روی 1 تنظیم می کنیم.


 کیفیت اندازه گیری کنترل را تعیین می کند. پارامترهای اندازه گیری کنترل می تواند تابعی از نوع و مکان گونه باشد، بنابراین اقدامات کنترلی بسیار خاص را می توان ایجاد کرد. برای واکسیناسیون، ما فرض می کنیم که زیست شناسی اساسی عامل اثربخشی غالب است، بنابراین این متغیرها وابسته به مکان نیستند. ما فرض می کنیم که تمام حیوانات تعیین شده برای معدوم سازی معدوم خواهند شد، بنابراین اثربخشی را روی 1 تنظیم می کنیم.
کیفیت اندازه گیری کنترل را تعیین می کند. پارامترهای اندازه گیری کنترل می تواند تابعی از نوع و مکان گونه باشد، بنابراین اقدامات کنترلی بسیار خاص را می توان ایجاد کرد. برای واکسیناسیون، ما فرض می کنیم که زیست شناسی اساسی عامل اثربخشی غالب است، بنابراین این متغیرها وابسته به مکان نیستند. ما فرض می کنیم که تمام حیوانات تعیین شده برای معدوم سازی معدوم خواهند شد، بنابراین اثربخشی را روی 1 تنظیم می کنیم.2.1. شبیه سازی ها
شبیهسازیهای ما به شکل زیر ساخته شدهاند: سطح بالای شبیهسازی، مدل بین شهرستانی است که دارای گام زمانی Δt = 1 روز است. در طول هر مرحله زمانی، همه کانتیها حلقهای هستند. اگر شهرستانی حیواناتی را آلوده کرده باشد، متغیرهای حالت { , , , , , , , , } با ادغام معادله (S3a-j) مدل درون شهرستانی با استفاده از روش Runge-Kutta مرتبه چهارم به روز می شوند. اگر یک شهرستان حیوانات آلوده نداشته باشد، معادله (1) به روز می شود و یک عدد تصادفی n∈ [0,1] از توزیع یکنواخت گرفته می شود. اگر n ≤ p X (t) سپس شهرستان آلوده خواهد شد، اگر n > p X (t) سپس شهرستان سالم باقی می ماند. در صورت آلوده شدن، حدود 100 حیوان بدون علامت آلوده معرفی میشوند و به حالت ناقل یا بدون علائم بالینی اختصاص داده میشوند، اما با توجه به نسبت انشعاب به حالت علائم پیشرفت میکنند . تعداد حیواناتی که در ابتدا آلوده شده بودند مانند Manore و همکاران انتخاب شد. [ 25 ]، تقریبی از تعداد حیوانات آلوده موجود قبل از شیوع بیماری تشخیص داده می شود، اما در صورت لزوم می توان آن را تغییر داد. ما مدل ریاضی دو سطحی بیماری همهگیر را برای شبیهسازی گسترش بیماری در میان جمعیت میزبان حساس چند گونه برای دو پاتوژن، FMD و HPAI به کار بردیم.
n > p X (t) سپس شهرستان سالم باقی می ماند. در صورت آلوده شدن، حدود 100 حیوان بدون علامت آلوده معرفی میشوند و به حالت ناقل یا بدون علائم بالینی اختصاص داده میشوند، اما با توجه به نسبت انشعاب به حالت علائم پیشرفت میکنند . تعداد حیواناتی که در ابتدا آلوده شده بودند مانند Manore و همکاران انتخاب شد. [ 25 ]، تقریبی از تعداد حیوانات آلوده موجود قبل از شیوع بیماری تشخیص داده می شود، اما در صورت لزوم می توان آن را تغییر داد. ما مدل ریاضی دو سطحی بیماری همهگیر را برای شبیهسازی گسترش بیماری در میان جمعیت میزبان حساس چند گونه برای دو پاتوژن، FMD و HPAI به کار بردیم. 
هر دو مجموعه شبیهسازی تنها با استفاده از دادههای سرشماری انبوه در سطح شهرستان ساخته شدند [ 24 ]. تکهها بهعنوان شهرستانهای منفرد ایالات متحده با کرسیهای شهرستان بهعنوان مرکز جغرافیایی تعریف میشوند. برای هر بیماری، ما چندین نمونه شبیهسازی را از توزیعهای پارامتری مناسب برای سه مکان مختلف شروع شیوع اولیه در کالیفرنیا، آرکانزاس و جورجیا برای HPAI و کالیفرنیا، آیووا و تگزاس برای FMD اجرا کردیم. برای هر مکان، ما 400 تحقق را اجرا کردیم، نمونه برداری از توابع چگالی احتمال پارامتر برای هر شرایط اولیه، با قطع زمانی 1000 روز.
سپس پیامد این دو بیماری را تعیین کردیم که به عنوان تعداد کل حیوانات آلوده در هر مرحله به عنوان تابعی از پارامترهای بیولوژیکی (مانند زمان کمون، دورههای عفونی علامتدار) و پارامترهای اندازهگیری کنترل (قرنطینه، کنترل حرکت، اثربخشی واکسیناسیون) اندازهگیری شد. . علاوه بر این، ما تأثیر جغرافیا (به عنوان مثال ، شرایط اولیه متفاوت) را بر گسترش همه گیری بررسی کردیم. در دو بخش فرعی بعدی، ما در مورد انتخاب های خاصی که برای پارامترهای مدل FMD و HPAI انجام شده است صحبت می کنیم.
2.2. مرض پا و دهان
ویروس FMD عضوی از خانواده ویروسهای Picornaviridae است که صمغها یا «سوکهای سم» مانند گاو، خوک، گوسفند، بز و گوزن را آلوده میکند [ 21 ]. بیماری بالینی ناشی از ویروس FMD با تب، بی اشتهایی و ظاهر شدن وزیکول در مخاط دهان و پا مشخص می شود. اگرچه مرگ و میر مرتبط با FMD معمولاً زیر 5 درصد است، عوارض می تواند به 100٪ برسد [ 26 ]. این بیماری با کاهش وزن و تولید شیر، ارزش تجاری حیوان را کاهش می دهد. FMD بیماری مشترک انسان و دام نیست، اگرچه انسان ها می توانند ویروس را به راحتی به مکان ها و حیوانات جدید منتقل کرده و پخش کنند [ 22 ]]. FMD بهره وری دام را تا حد زیادی کاهش می دهد و کشورهای آسیب دیده نمی توانند در تجارت بین المللی حیوانات و محصولات حیوانی شرکت کنند.
گاوها واضح ترین علائم بالینی را نشان می دهند و بنابراین می توان آنها را حیوان نگهبان در نظر گرفت. گوسفند و بز علائم بالینی قابل توجهی نشان نمی دهند و می توانند به راحتی بیماری را به سایر حیوانات اهلی منتقل کنند [ 27 ، 28 ، 29 ، 30 ، 31 ]. روش های اولیه انتقال FMD عبارتند از: آئروسل ها، تماس مستقیم و غیرمستقیم با حیوان آلوده و بلعیدن. حرکت حیوانات آلوده تا حد زیادی مهم ترین عامل انتقال فضایی است و به دنبال آن جابجایی محصولات حیوانی آلوده [ 32 ].
ما از دادههای مکانی آمار کشاورزی وزارت کشاورزی ایالات متحده در سال 2007 [ 24 ] برای تنظیم اولیه توزیع جمعیت در سطح شهرستان از گونههای مورد علاقه، به عنوان مثال ، گاو گوشتی، گاو شیری، گاو در پروار، خوک و گوسفند استفاده کردیم. شکل 2 a,b تراکم گراز و گاو را به ترتیب در ایالات متحده از سرشماری سال 2007 نشان می دهد. در جدول 1، پارامترهای مدلی را که در شبیه سازی استفاده کرده ایم نشان می دهیم. ما همه پارامترهای بیولوژیکی و اپیدمیولوژیک را به طور معمولی توزیع کردیم. مقادیر میانگین و انحراف استاندارد برای توزیع پارامترها (مگر اینکه به صراحت در مرجع ذکر شده باشد) از منابع ادبیات با در نظر گرفتن میانگین به عنوان مرکز دامنه مقادیر مشخص شده تعیین شد و انحراف استاندارد نیمی از محدوده در نظر گرفته شد. پارامترهای اندازه گیری کنترل به طور یکنواخت در محدوده های مشخص شده خود توزیع می شوند. مقادیر شروع برای پارامترهای انتقال و حساسیت برای گاوهای علامت دار از تلاش های قبلی مدل سازی FMD [ 23 ] گرفته شد.]. ما این فرض را داریم که حیوانات در حالت ناقل بدون علائم بالینی، نصف حیوان علامت دار عفونی هستند، زیرا عفونی بودن حالت ناقل بسیار نامشخص است [ 33 ]. هنگامی که گرازها و خوک ها علائم بالینی را نشان نمی دهند، عفونتی مشابه گاو یا گوسفند دارند، اما عفونی بودن آنها نسبت به گاو در مرحله علامتی دو برابر افزایش می یابد، زیرا تمایل آنها به تولید قطرات آئروسل حاوی غلظت های بالایی از این ماده است. ویروس [ 34 ]. ما پارامترهای انتقال را برای گوسفندان یکسان با گاو تنظیم کردیم. مقیاس طول کوتاه گسترش در پنج مایل تنظیم شده است، که تا حدودی بزرگتر از هسته به دست آمده تجربی است، تخمین زده شده توسط DEFRA، در طول شیوع سال 2001 انگلستان (4 کیلومتر) [ 23 ]]، که مکانیسم های رایج حرکت حیوانات و انتقال ویروس توسط کامیون ها، تجهیزات و فومیته ها بود. برای گاو، گراز و گوسفند، داده های مفصلی در مورد حرکات بین ایالتی حیوانات وجود دارد [ 35 ]. ما داده های ماهانه را به فرکانس برای مدت حمل و نقل حیوانات از راه دور تبدیل کردیم.
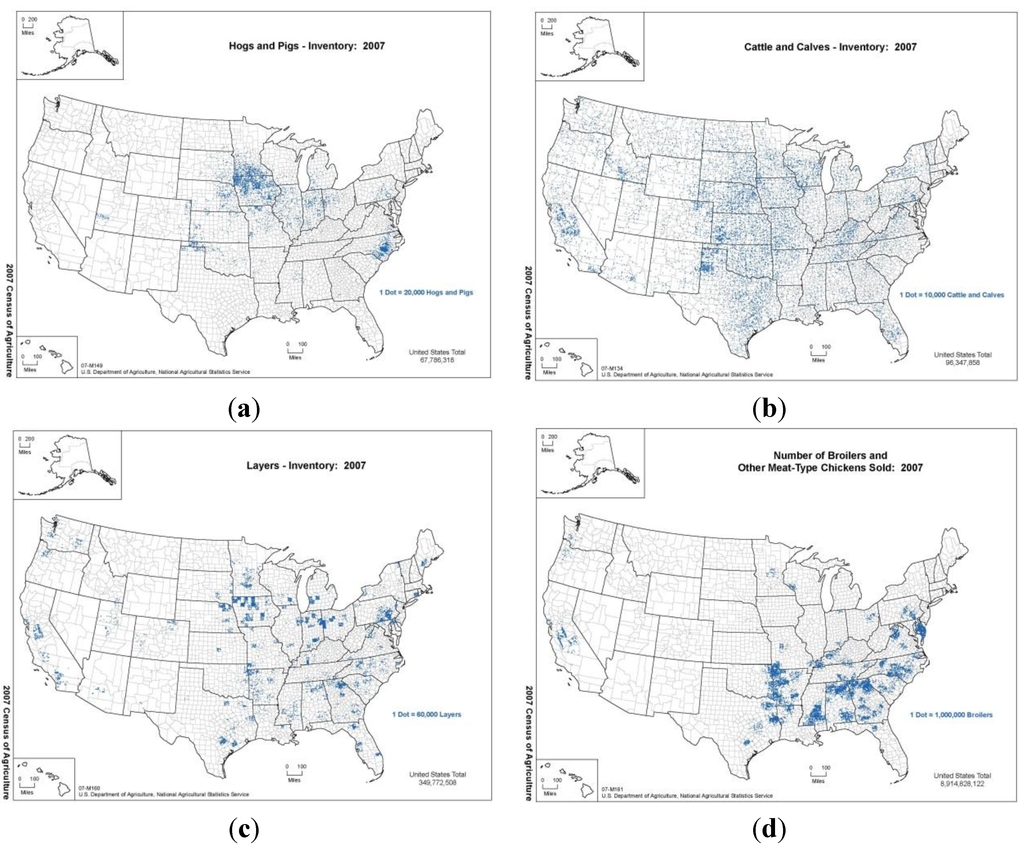
شکل 2. نمودارهای تراکم ( الف ) گراز و خوک، ( ب ) گاو و گوساله در ایالات متحده بر اساس سرشماری USDA NASS 2007 (ردیف بالا، چپ به راست). نمودارهای تراکم ( c ) لایه ها و ( d ) جوجه های گوشتی در ایالات متحده بر اساس سرشماری USDA NASS 2007 (ردیف پایین، چپ به راست). در شبیهسازیهای ما، مناطق با تراکم بالای گونههای میزبان منجر به گسترش سریعتر و پیامدهای بالاتر میشوند.

جدول 1. پارامترهای مدل اندازه گیری بیولوژیکی و کنترلی مورد استفاده برای شبیه سازی بیماری تب برفکی (FMD).
پارامترهای پیشرفت بیماری از منابع مختلف ادبیات گرفته شد. زمان جوجه کشی برای گاو معمولاً 4 تا 8 روز است [ 36 ، 37 ]. خوک ها به طور کلی سریعتر پیشرفت می کنند و می توانند علائم را 2 تا 5 روز پس از عفونت نشان دهند [ 27 ، 38 ]. گوسفندها معمولاً به مدت 14 تا 21 روز پیشرفت می کنند [ 39 ]. بیشتر پراکندگی در زمان جوجه کشی نشان دهنده عدم قطعیت در حالت انتقال و اندازه دوز تلقیح است. مرحله علامتی برای همه گونه های جانوری نسبتا یکنواخت است و 14 تا 21 روز طول می کشد [ 32 , 40 ]]. حیوانات ناقل می توانند در هر دو حالت علائم بالینی و غیر بالینی برای مدت طولانی عفونی باقی بمانند، به عنوان مثال، 4 ماه تا 3.5 سال برای گاو. در حالی که 95٪ تا 100٪ از همه گاوها و گرازها به علائم بالینی پیشرفت می کنند [ 32 ، 41 ]، گوسفندها به ویژه برای پیشرفت به حالت ناقل علائم غیر بالینی حساس هستند [ 30 ، 42 ]. تا 25% از گوسفندان آلوده دچار ضایعات نمی شوند، بنابراین ما محدوده وسیعی (25% تا 50%) را برای بخشی از گوسفندان آلوده که به علائم پیشرفت می کنند اختصاص می دهیم.
مدت زمانی که حیوان پس از پاکسازی ویروس مصونیت دارد، متناسب با زمان پوسیدگی آنتی بادی های خنثی کننده ناشی از عفونت اولیه است. تنوع آنتی ژنی FMD به خوبی شناخته شده است و عفونت با یک سروتیپ FMD محافظتی در برابر سایر زیرگروه ها ایجاد نمی کند [ 39 ، 43 ]. بنابراین، مصونیت در برابر عفونت مجدد با زیرگروه یکسان دائمی نیست و طبق گفته Carrillo و همکاران ، تقریباً 90 تا 180 روز برای گاو، گراز و گوسفند است. [ 36 ] و اورسل و همکاران. [ 37 ].
مفروضات زیر برای پارامترهای نظارت و پاسخ نشان داده شده در جدول 1 انجام شد. در سطح شهرستان، ما یک فاصله زمانی 1 تا 21 روزه بین تشخیص و شروع هر دو واکنش قرنطینه و کشتار داریم. یک محدوده توزیع یکنواخت از (0.10-0.90) برای اثربخشی کنترلهای حرکتی برد کوتاه و بین ایالتی و همچنین قرنطینه انتخاب شده است تا درجه بالایی از عدم قطعیت را در این پارامترها منعکس کند. با توجه به اینکه آخرین شیوع بیماری در ایالات متحده در سال 1929 بود، عدم اطمینان زیادی در عواملی مانند انطباق و سطح منابع متعهد وجود دارد که تا حدی به دلیل عدم تجربه عملی در مورد FMD است. یک واکسن همسان FMD برای درمان و پیشگیری تا 17 روز پس از تشخیص در شبیهسازیهای ما در دسترس نیست، که نشاندهنده زمان مورد نیاز برای مطابقت با ژنوتیپ و تولید انبوه است.
2.3. آنفولانزای فوق حاد پرندگان
آنفولانزای پرندگان یک بیماری عفونی جهانی پرندگان است که توسط سویههای نوع A ویروس آنفلوانزا ایجاد میشود. بسیاری از گونههای پرندگان وحشی حامل این ویروسها هستند و هیچ نشانهای از آسیب ندارند. در طیور، ویروسها باعث ایجاد دو نوع متفاوت از بیماری میشوند-یکی شایع و خفیف (آنفولانزای پرندگان با بیماریزایی کم)، دیگری نادر و بسیار کشنده (HPAI) با نرخ تلفات که میتواند به 100 درصد برسد. علائم بالینی ممکن است شامل ضایعات، افسردگی شدید، از دست دادن اشتها، کاهش تولید تخمک، ادم صورت، و خونریزی در سطوح غشای داخلی باشد. پرندگان آلوده ویروس را از طریق ترشحات مدفوع و بینی دفع می کنند. ویروس آنفلوانزای مرغی جدا از اینکه در بین طیور بسیار مسری است، میتواند از طریق جابجایی طیور زنده، افراد (به ویژه کفش و سایر لباسها)، وسایل نقلیه، تجهیزات، خوراک و قفس بین مزارع منتقل شود. علاوه بر این، بیماری می تواند از طریق دفع نامناسب لاشه آلوده، کود دامی یا محصولات جانبی طیور سرایت کند. حشرات و جوندگان ممکن است به طور مکانیکی ویروس را از طیور آلوده به طیور حساس منتقل کنند.
ما مدل را با دادههای سرشماری طیور در سال 2007 وزارت کشاورزی ایالات متحده، خدمات آماری ملی کشاورزی [ 24 ] در مورد لایهها، پولتها، جوجههای گوشتی و بوقلمونها تکمیل کردیم. شکل 2 c,d تراکم لایه ها و جوجه های گوشتی را به ترتیب در ایالات متحده از سرشماری سال 2007 نشان می دهد. جدول 2 پارامترهای مدل را برای شبیه سازی HPAI نشان می دهد. همانطور که در مورد FMD، ما فرض می کنیم که پارامترهای بیولوژیکی/اپیدمیولوژیک به طور معمول توزیع شده اند در حالی که پارامترهای اندازه گیری کنترل به طور یکنواخت توزیع شده اند. پارامترهای مدل انتقال داخل شهرستان برای بازتولید نرخ حمله گله 60٪ مشاهده شده در مرغداری ها در طول رویدادهای اپیزوتیک HPAI آسیایی کالیبره شدند [ 43 ]] در اجراهای جداگانه با اقدامات کنترلی تنظیم شده برای مطابقت با آن شیوع.
مکانیسم های انتشار بیماری شامل تماس پرنده به پرنده در سطح مزرعه فردی، جابجایی حیوانات آلوده، و راه های انتقال با واسطه انسان شامل کامیون ها، تجهیزات یا سایر مواد آلوده است. ما فرض می کنیم که هیچ یک از پرندگان تا زمانی که علائمی را نشان ندهند، عفونی نباشند، که با مسیر مدفوع به دهان مطابقت دارد که در آن ویروس باید در هنگام اسهال به مکان هایی که ممکن است پرندگان نوک بزنند، ریخته شود. فقط پراکندگی محلی بین تأسیسات شبیه سازی شد، با این فرض که پرندگان به طور متوسط در فاصله تقریباً پنج مایلی بین جایی که از تخم بیرون می آیند و جایی که ذبح می شوند (تولید) ظاهر می شوند، که با مشاهده این که حمل و نقل بین ایالتی طیور مطابقت دارد. محدود [ 44 ].
جدول 2. پارامترهای مدل اندازه گیری بیولوژیکی و کنترلی مورد استفاده برای شبیه سازی آنفولانزای پرندگان بسیار بیماری زا (HPAI).
محدوده پارامترهای پیشرفت بیماری بر اساس مقادیر ادبیات گزارش شده برای شیوع اخیر HPAI بود. دوره جوجه کشی در طیور می تواند بین 1.6-8 روز بسته به سویه متفاوت باشد [ 45 ]. ما محدوده ای را در زمان های جوجه کشی که توسط کار Shortridge و همکاران ارائه شده بود، انتخاب کردیم. [ 46 ]. برای دوره عفونی (و با فرض اینکه ریزش ویروس در طیور ساده لوح زیاد خواهد بود) محدوده 1-1.5 روز را انتخاب کردیم [ 45 ]. پس از این مدت، طیور با نرخ مرگ و میر بالینی 97.5 درصد بهبود می یابند یا می میرند. ما مصونیت طیور بهبودیافته را در برابر عفونت های مکرر فرض می کنیم [ 47 ، 48 ، 49 ، 50 ].
استراتژی های کاهش ارزیابی شده قرنطینه، حذف و کنترل حرکت است. واکسیناسیون انتخاب نشد زیرا می تواند منجر به ریزش ویروس در پرندگان واکسینه شده و غیرآلوده شود، و همچنین، ایجاد علائم غیر بالینی “حامل”، که می تواند خطر انتقال بین گله را قبل از تشخیص همه گیری افزایش دهد [ 51،52 ، 53 ]. پارامترهای مورد استفاده در شبیه سازی ها در جدول 2 نشان داده شده است.
اثربخشی کلی کنترلهای کاهش و حرکت، هم در مقیاس منطقهای و هم در مقیاس ملی (بین ایالتی) در طول یک شیوع بزرگ HPAI، تا حد زیادی نامشخص است. تجارب گذشته در ایالات متحده با HPAI (پنسیلوانیا 1983، تگزاس 2004) نشان می دهد که واکنش سریع و اقدامات امنیت زیستی خوب مهم هستند. حداکثر اثربخشی کنترلهای حرکتی بین شهرستانی به عنوان نسبتاً کم (50٪) اختصاص داده شده است و کارایی قرنطینه به دلیل پدیدهای به نام گسترش منطقه [ 54 ] شناخته میشود که شامل طیف کامل عدم قطعیت ( یعنی 0-1) میشود. نقض ناشناخته امنیت زیستی و سایر شیوه های سنتی مزرعه و جامعه که می تواند منجر به جابجایی عوامل عفونی شود. این امر برای HPAI که در محیط ایجاد می شود از اهمیت ویژه ای برخوردار است [ 54, 55 , 56 ].
برای هر بیماری، HPAI و FMD، ما 400 اپیدمی جداگانه را شبیهسازی کردیم که در هر یک از سه مکان شروع شد. برای هر اجرای شبیهسازی، 100 حیوان در جمعیتی کاملاً متشکل از حیوانات حساس در شروع روز 1 با پارامترهای پیشرفت بیماری و اثرات کاهشدهنده بهطور تصادفی از محدودههای ارائهشده در جدول 1 (برای HPAI) و بهطور تصادفی انتخاب شدهاند. جدول 2 (برای FMD). اپیدمی به طور تصادفی شبیه سازی شد و یک فایل خروجی حاوی جمعیت هر نوع حیوان در هر شهرستان ایالات متحده برای هر مرحله بیماری تولید کرد ( شکل 1 را ببینید.) در هر روز اپیدمی. شبیهسازیهای اپیدمی در 1000 روز پایان یافتند، اگر قبلاً خاموش نشده بودند، و اعداد کل پیامدهای ارائهشده در اینجا فقط شامل 1000 روز اول برای این همهگیریها میشود.
برای هر بیماری، ما توزیع حیوانات را در سراسر ایالات متحده بررسی کردیم و مکانهایی را انتخاب کردیم که خوشههای متمایز از دام را نشان میداد. برای HPAI، ما اپیدمی را در کالیفرنیا، آرکانزاس، و جورجیا به دلیل تنوع جغرافیایی نشاندهنده هر یک از ایالتها و تراکم بالای لایهها و جوجههای گوشتی در این ایالتها آغاز کردیم. برای FMD، ما کالیفرنیا، تگزاس و آیووا را انتخاب کردیم تا تنوع جغرافیایی را در نظر بگیریم و در عین حال ایالتهایی با تراکم گاو (CA، TX) و خوک (IA) را در نظر بگیریم. در در نظر گرفتن تأثیر جغرافیا بر گسترش اپیدمی، توجه به این نکته مهم است که ما یک حمل و نقل بین ایالتی حیوانات را در مدل FMD، بر اساس تحرکات دام بین ایالتی منتشر شده وزارت کشاورزی ایالات متحده [ 57 ]، در حالی که فرض می کنیم حمل و نقل بین ایالتی طیور وجود ندارد، گنجانده ایم. رخ داد (شکل 2 ). این تا حدی به دلیل ماهیت صنایع و تا حدی به دلیل افزایش توانایی جداسازی طیور از حیات وحش در مقایسه با گاو است.
ما از روشهای انتخاب مدل آماری (AIC) برای شناسایی محرکهای اصلی شدت یا پیامدهای اپیدمی در بین پارامترهای مدل بیماری و کنترل استفاده میکنیم [ 58 ]. پارامترهای بیماری که بهعنوان قابلتوجه شناخته میشوند، نشان میدهند که کدام عناصر زیستشناسی میزبان-پاتوژن و استراتژیهای کاهش اندازه اپیدمی را هدایت میکنند. به همین ترتیب، ما تشخیص میدهیم که اقدامات کنترلی برای کنترل شیوع بیماری و وابستگی به اثربخشی این استراتژیهای کاهش حیاتی هستند.
3. نتایج
دو عامل بزرگ برای هزینه اقتصادی کلی اپیدمی های دام، بزرگی و مدت اپیدمی است، و ما روز اوج اپیدمی و کل حیوانات را برای HPAI و FMD مقایسه می کنیم ( شکل 3 ). هر نماد خروجی کلی یک اپیدمی شبیهسازی شده را نشان میدهد، با اندازه کلی اپیدمی، که در تعداد کل حیوانات آلوده اندازهگیری میشود و روی محور y لگاریتمی ترسیم میشود، و روز اوج همهگیری در محور x. برای هر دو بیماری، جغرافیا پیشبینیکننده خوبی برای سرعت و مقیاس همهگیری بود. شکل 4نمایش نماینده ای برای HPAI با سه مکان شروع را نشان می دهد. برای HPAI، اپیدمیهای آغاز شده در کالیفرنیا کوچک باقی میمانند و به سرعت به اوج میرسند، در حالی که اپیدمیهای آغاز شده در آرکانزاس به سرعت به اوج خود میرسند، اما به طور کلی، بزرگترین اپیدمی از این سه مکان هستند. اپیدمی ها در جورجیا شروع شدند، در حالی که تقریباً به اندازه موارد شروع شده در آرکانزاس بودند، به جای 1 هفته مشخصه موارد شروع شده در کالیفرنیا و 10 روز مشخصه موارد شروع شده در آرکانزاس، تنها پس از 3 تا 6 هفته به اوج خود می رسند.
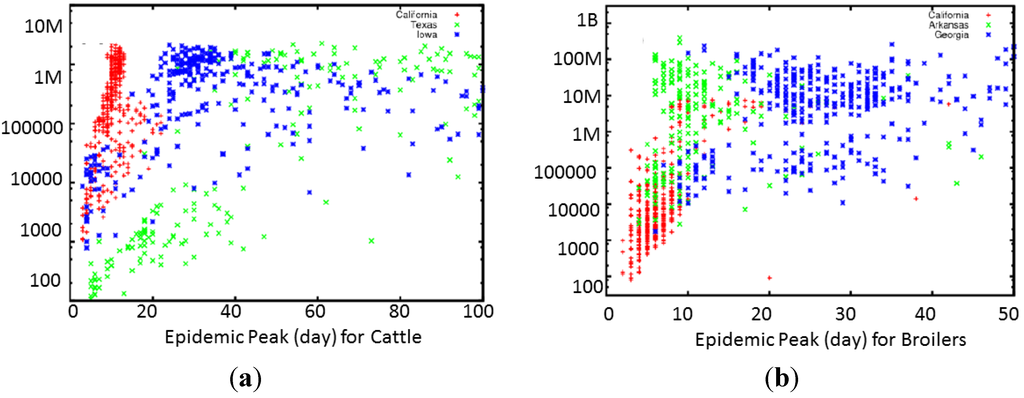
شکل 3. نمودار پراکندگی تعداد کل حیوانات آلوده در برابر طول مدت اپیدمی برای 1200 تحقق مدل پیامد برای ( الف ) FMD و ( ب ) HPAI. هر نقطه مربوط به یک اجرا با مجموعه ای از پارامترهای نمونه برداری تصادفی است. رنگ های مختلف محل اولیه گسترش همه گیری را نشان می دهد. اگرچه هر دو نمودار وابستگی به جغرافیا را نشان میدهند، نمودار HPAI بسته به محل شروع همهگیری، خوشهبندی بسیار دقیقتری از طول و اندازه اپیدمی نشان میدهد. یک استثنا این است که برای FMD، شیوع بیماری در کالیفرنیا به اوج و سریع می رسد.

شکل 4. گسترش سطح بین شهرستانی HPAI. طبق دادههای سرشماری کشاورزی USDA NASS در سال 2007 برای لایهها، پولتها، جوجههای گوشتی و بوقلمونها، نقاط سبز نشاندهنده شهرستانهایی است که در آن طیور مستعد وجود دارد. نقاط آبی نشاندهنده شهرستانهایی است که حداقل 10 پرنده بدون علامت آلوده وجود دارد، نقاط قرمز نشاندهنده شهرستانهایی هستند که حداقل 1 پرنده آلوده علامتدار دارند. صلیبهای سیاه نشاندهنده شهرستانهایی هستند که یا در ابتدا هیچ مرغ حساسی ندارند یا در آنها افراد مستعد از طریق اقدامات قرنطینه، حذف یا تلفات ناشی از بیماری تخلیه شدهاند.
برای اپیدمی های HPAI، اثر زمان نهفتگی بر سرعت کلی اوج گیری اپیدمی زیاد است، با زمان های نهفتگی کوتاه که منجر به پیامدهای بالاتر و پیک های اپیدمی بسیار سریع می شود. این منعکس کننده تصمیم ما برای نمونه برداری از زمان های جوجه کشی از 1 تا 7 روز است که با تنوع گسترده مشاهده شده در این پارامتر مطابقت دارد و ماهیت نسبتاً ایزوله کالیفرنیا شرایط را برای شبیه سازی اپیدمی بسیار تکرارپذیر فراهم می کند.
جدول 3. خروجی های انتخاب مدل (AIC) برای آنفولانزای پرندگان بسیار بیماری زا.
برای نقاط پایانی اپیدمی برای HPAI، زمان اوج اپیدمی یا سرعت گسترش آن تحت تأثیر نرخ مرگ و میر بیماری ذاتی (ANOVA، F 11194 = 0.90، P = 0.34) یا روزی که پاسخ ایجاد شد (F 1,1194 = 3.20، P = 0.075). تعداد کل حیوانات مرده به روزی که پاسخ ایجاد شد نیز وابسته نبود (F 1,1196 = 1.93، P = 0.165). تعداد کل حیوانات آلوده نیز تحت تأثیر نرخ مرگ و میر ذاتی قرار نگرفت (F 11194 = 0.24، P = 0.62). تمام ویژگی های دیگر بیماری و کاهش آن که متفاوت بود بر شدت و مدت اپیدمی HPAI تأثیر گذاشت.جدول 3 ).
در مقابل، برای FMD، همبستگی بسیار کمی بین زمان نهفتگی و مقیاس یا مدت اپیدمی وجود دارد. این احتمالاً به دلیل تنوع نسبتاً کم مشاهده شده در این پارامتر است، و به این دلیل که جنبه حمل و نقل بین حالتی مدل ما نویز تصادفی قابلتوجهی را به پیامد کلی شبیهسازی معرفی میکند. شکل 5 مقیاس در مقابل سرعت را برای یک تحقق واحد از شبیه سازی FMD برای گاوهای گوشتی، گرازها و گاوهای شیری برای یک تحقق نشان می دهد. مانند HPAI، جغرافیا در گسترش FMD مهم است. با این حال، برخلاف HPAI، با حمل و نقل دام از راه دور و موارد تحت بالینی بیشتر، شیوع FMD اغلب به سرعت به سطوح ملی افزایش یافت. شکل 6یک اجرای معمولی برای FMD را نشان می دهد که در سه شهرستان شروع شده است (پانل بالا سمت چپ).
شکل 7 و شکل 8 حساسیت بزرگی پیامد به اقدامات کنترلی را نشان می دهد. در شکل 7 ، نشان میدهیم که چگونه میزان شیوع FMD (اندازهگیری شده بر حسب گاو مرده گوشت گاو) با تأخیر زمانی بین تشخیص بیماری و شروع معدوم کردن حیوانات آلوده متفاوت است. شکل 8 نشان می دهد که چگونه اثر قرنطینه در کاهش پیامد HPAI از نظر کل طیور مرده همبستگی دارد.

شکل 5. سری های زمانی (میانگین در شهرستان ها و شبیه سازی ها) برای تعداد حیوانات تازه آلوده از FMD در برابر زمان اوج اپیدمی ترسیم شده است، با رنگ نشان دهنده نوع حیوان. به طور متوسط، شیوع FMD در حدود 80 روز به اوج خود می رسد و گرازها بیشترین آسیب را دارند. اپیدمی در گاوهای گوشتی با تعداد کمتری به اوج خود می رسد، اما بیشتر از گراز طول می کشد.
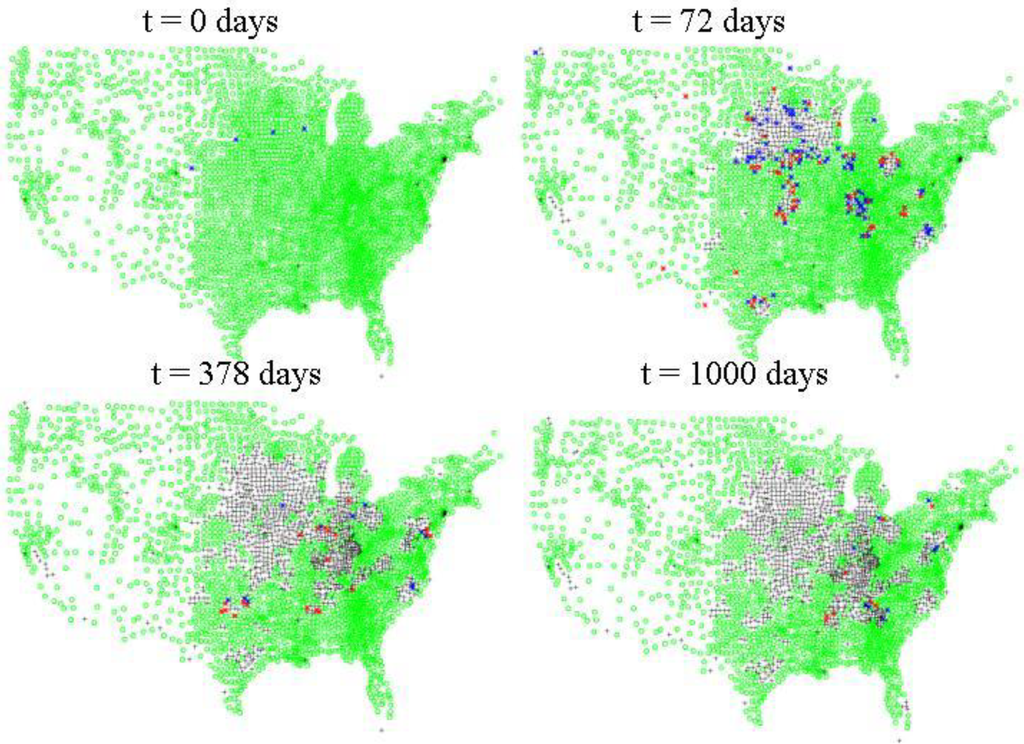
شکل 6. گسترش سطح بین شهرستانی FMD. طبق دادههای سرشماری کشاورزی 2007 USDA NASS، نقاط سبز نشان میدهد که در آن جمعیتهای مستعد گاو، گراز و/یا گوسفند وجود دارد. نقاط آبی نشان می دهد که در آن 10 حیوان بدون علامت یا بیشتر وجود دارد، نقاط قرمز نشان می دهد که یک یا چند حیوان علامت دار وجود دارد. صلیبهای سیاه نشاندهنده شهرستانهایی هستند که یا هیچ جمعیت اولیه حساسی نداشتند یا با اقدامات کاهشدهنده، یعنی قرنطینه، حذف و/یا واکسیناسیون، از جمعیت مستعد خالی شدهاند .

شکل 7. سری های زمانی برای همه شبیه سازی ها برای FMD گروه بندی شده بر اساس زمان کشتن حیوانات. نشانههای قرمز بعلاوه، تاخیر جمعآوری 14 تا 21 روز، رنگ سبز با تأخیر جمعآوری 7 تا 14 روز و ستارههای آبی تاخیر جمعآوری 1 تا 7 روز است. تأخیر کمتر در معدوم سازی عموماً منجر به کاهش کل گاوهای مرده و انتشار آهسته تر اپیدمی می شود.
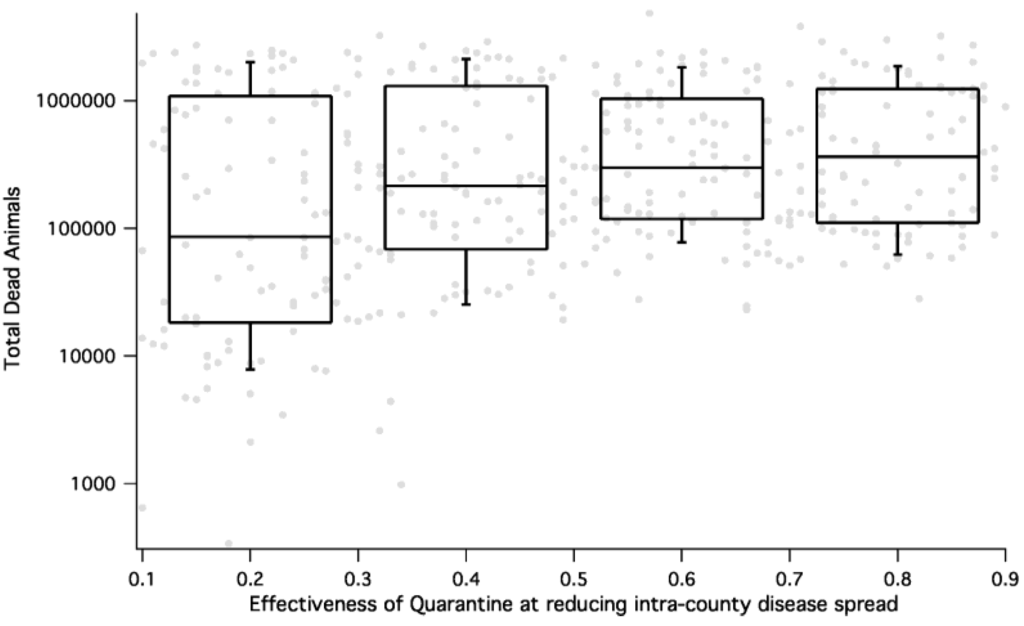
شکل 8. پیامد HPAI (به عنوان تعداد کل مردگان در طول شبیه سازی اندازه گیری می شود) به عنوان تابعی از اثربخشی قرنطینه برای همه شبیه سازی های همه گیر. درصد حیواناتی که توسط قرنطینه محافظت نمی شوند در محور x و تعداد کل حیوانات مرده در محور y است. ما می بینیم که برای HPAI، قرنطینه موثر منجر به کاهش حیوانات مرده تقریباً یک مرتبه می شود.
4. بحث
ما یک چارچوب مدل با دانه بندی مناسب (با توجه به داده های محدود در مورد موقعیت جغرافیایی مزارع دام و حرکت حیوانات) و پیچیدگی برای شبیه سازی انواع بیماری های عفونی با کاهش ارائه کردیم و استفاده از آن را در تجزیه و تحلیل شیوع HPAI و FMD نشان دادیم. این مدل امکان توصیف واقعی تفاوتها در پیشرفت بیماری و توزیع جغرافیایی انواع میزبانهای مختلف، و همچنین تفاوتها در توانایی ما برای کاهش شیوع را فراهم میکند. این مدل همچنین با سرعت کافی اجرا می شود که حساسیت پیامد به پارامترهای پیشرفت بیماری، پارامترهای کاهش بیماری و جغرافیا را می توان مشخص کرد. با این حال، نیازمند این است که محدوده های واقعی برای همه پارامترها به طور مناسب تعریف شود تا این تحلیل حساسیت به درستی کار کند.
جغرافیا یک جنبه مهم از همه گیری است که توسط مدل ما گرفته شده است. در واقع، گسترش HPAI ویژگیهای انتشار مانندی را در چارچوب موانع جغرافیایی و تراکم میزبان متفاوت در میان شهرستانها نشان میدهد. این نوع گسترش فضایی به راحتی با انزوا یا قرنطینه کاهش می یابد همانطور که در شکل 8 مشاهده می شود.و ممکن است با توجه به زمان قبل از شروع تخفیف، بخشنده تر باشد. همانطور که انتظار می رود، FMD، که به سرعت در سراسر کشور گسترش می یابد، به نظر می رسد که جنبه های عمده ای از ویژگی های همه گیر آن توسط توزیع فضایی حیوانات میزبان و حامل تعیین می شود. اگرچه هیچ تفاوتی در پیامدهای اپیدمی برای FMD بر اساس مکان های شروع انتخابی ما وجود نداشت، این مورد برای HPAI در طیور صادق نبود. با این گفته، ارزش بررسی ساختار هسته گسترش وابسته به فاصله و ماتریس حمل و نقل دوربرد ما را دارد. به طور خاص، هنگام مقایسه نتیجه در مقابل.جغرافیای شروع، اپیدمی هایی که در کالیفرنیا شروع می شوند، منعکس کننده وابستگی به مکانیسم های حمل و نقل دوربرد هستند. اپیدمی هایی که در کالیفرنیا شروع شده اند معمولاً یک نوار محکم را تشکیل می دهند، زیرا یک سیستم منزوی و همگن است. پراکندگی بزرگتر در مناطق با تراکم میزبان بالا و اتصال بالا اتفاق می افتد (به عنوان مثال، آرکانزاس و جورجیا برای HPAI و غرب میانه برای FMD).
ما تصدیق میکنیم که ضعف شبیهسازیهای نشاندادهشده در اینجا این است که شهرستان احتمالاً بهترین واحد اپیدمیولوژیک برای گسترش بیماریهای حیوانی نیست، و احتمالاً برای دقت بالا به وضوح در سطح مزرعه نیاز است [ 59 ]]. با این حال، ما نشان میدهیم که یک مدل به درستی فرمولبندی شده که حاوی تمام ویژگیهای بارز انتقال بیماری است، میتواند نتایج معقولی ارائه دهد و حساسیتهای صحیح را به توزیعهای میزبان جغرافیایی و اقدامات کنترلی در غیاب دادههای با وضوح بالاتر ارائه دهد. اگر دادههای سطح مزرعه در دسترس باشد، این چارچوب مدل میتواند به گونهای تطبیق داده شود که گرهها مزارع را به جای شهرستانها نشان دهند. ما انتظار داریم که در این ساختار، گسترش درون شهرستانی کند شود، و استنباط کنیم که شبیهسازیهای نشاندادهشده در اینجا عموماً نمایانگر یک «بدترین سناریو» هستند و در عین حال بینشی در مقیاس گسترده برای کاهش و نظارت در سطح شهرستان یا ایالت ارائه میدهند.
چیزهای زیادی وجود دارد که بر گسترش اپیدمی بیماری عفونی در حیوانات تأثیر میگذارد که شامل ژنتیک میزبان [ 60 ] و محیط [ 61 ] میشود – با پیامدهای بالقوه برای تأثیرگذاری بر گسترش بیماری مرتبط با تغییرات آب و هوایی [ 62 ]. با این حال، استفاده از مدلهای مقیاسبندی مناسب برای مطابقت با دادههای جمعآوریشده از اپیدمیهای قبلی و ویژگیهای بیماری مشاهدهشده میتواند به کاهش اثرات این اثرات خرد بر شبیهسازیهای کلی کمک کند. علاوه بر این، استفاده از شبیهسازیهای متعدد بر اساس تمام محدودههای بالقوه پارامترهای بیماری، همانطور که در این مطالعه انجام شد، میتواند به کمیت کردن تغییراتی که ممکن است در هر شیوع آینده مورد انتظار باشد کمک کند.
کاربرد این مدل در سناریوهای FMD و HPAI توانایی مدل را در ضبط شیوع بیماریهای بسیار متفاوت با میزبانهای مختلف نشان میدهد. نقاط قوت این مدل شامل ادغام ماهیت جغرافیایی شبکه تماس میزبان بدون هیچ گونه کالیبراسیون بسیار خاص نسبت به یک بیماری خاص، سهم گونههای مختلف و انواع دام در اپیدمیولوژی، دامنه بزرگی علائم و غیره است . ناشی از تعامل میزبان و پاتوژن)، و واریانس در خود پارامترهای بیماری، که منعکس کننده عدم قطعیت در حدت یک پاتوژن خاص است. ادغام کاهش در مقیاس های چندگانه نیز سهم مهمی در حرکت به سمت مدل سازی برای سیاست است.
ذکر این نکته حائز اهمیت است که تشخیص بیماری به کشاورزان و ذینفعان بستگی دارد که نه تنها بیماری را شناسایی کنند، بلکه در مورد هر حیوان بیمار گزارش دهند. در حالی که گزارش رسمی بیماری در حیوانات کار درستی است، اما به دلیل پتانسیل هزینه های اقتصادی شدید، شجاعت می طلبد. این مهم است که به کشاورزان و عموم مردم آموزش داده شود که نه تنها ثابت شده است که تشخیص سریع بیماری باعث کندی و حتی جلوگیری از شیوع آن می شود، بلکه اقتصاد کلی به طور قابل توجهی کمتر از تشخیص بیماری در آینده است [ 63 ، 64 ].
این جنبه ها چارچوب مدل را برای ارزیابی تهدیدات بیماری های عفونی در حال ظهور ایده آل می کند. بیش از 70 درصد از مهم ترین عفونت های انسانی (ایدز، وبا، تب دنگی، SARS، آنفولانزای H5N1) منشأ مشترک بین انسان و دام دارند که از مخازن شناخته شده و ناشناخته حیات وحش ناشی می شوند. گله های حیوانات اهلی بزرگ از دام های کشاورزی می توانند مجرای کارآمدی برای انتقال پاتوژن از حیوانات وحشی به انسان فراهم کنند [ 1 ]]. حتی بدون سرایت به انسان، پاتوژن های موثر بر دام ها تهدیدی برای امنیت غذایی محسوب می شوند و می توانند خسارات اقتصادی قابل توجهی ایجاد کنند. نظارت ژنتیکی، محیطی یا اپیدمیولوژیک زودهنگام “نقاط داغ” بیماری یا شیوع اضطراری می تواند در ابتدا داده های پراکنده و/یا کم وفاداری به دست دهد. با توجه به اینکه شبیهسازیهای ما میتوانند از این عدم قطعیت نمونهبرداری کنند، این مدل میتواند تخمینهای اولیه و پیامدهای اولیه را تولید کند که میتواند به تدریج به طور مکرر اصلاح شود، به عنوان مثال، از طریق تکنیکهای تخمین بیزی همانطور که در [ 65 شرح داده شد.] با در دسترس قرار گرفتن داده های بیشتر. جمعیتهای انسانی، با جفتهای مناسب به جمعیتهای حیوانی، میتوانند برای ارزیابی خطر مشترک بین انسان و دام اضافه شوند. این دانش از برون یابی ساده بیماری های شناخته شده پشتیبانی تصمیم گیری موثرتری را برای فعالیت هایی مانند اولویت بندی تخصیص منابع و ایجاد یا اصلاح خط مشی ارائه می دهد.
این چارچوب مدل را می توان با مراحل شیوع چندگانه تطبیق داد. وصله ها یا گره های شبکه در مدل می توانند محدوده و تعریف را با پیشرفت شیوع تغییر دهند. در شروع شیوع، گره ها را می توان به عنوان میزبان فردی تعریف کرد. پارامترهای بیماری برای انتقال و پیشرفت را می توان از تراکم احتمال تخمین زده شده از داده های اولیه نمونه برداری کرد. در طول این مرحله، از مدل معادلات دیفرانسیل معمولی درون وصله استفاده نمی شود زیرا نوسانات آماری غالب هستند و تقریب اختلاط یکنواخت برای این مرحله از رویداد معتبر نیست. در نهایت، اگر دامنه شیوع افزایش یابد، تکهها یا گرهها برای نشان دادن واحدهای اپیدمیولوژیک مختلف در مقیاسهای مختلف (مثلاً گروههای خانواده، گلهها، مزارع، شهرها، شهرستانها و غیره ) تغییر میکنند.). تغییرات پیکربندی وصله عمدتاً به صحت دادههای موجود بستگی دارد، اما همچنین به ارزیابیهای کیفی مربوط به جایی که انتظار میرود اختلاط یکنواخت وجود داشته باشد یا اینکه کدام انتخاب واحد اپیدمیولوژیک نمایندگی پچ بهترین تناسب را برای تخمین پارامتر مدل میدهد، نیز بستگی دارد. ما انتظار داریم که این فرآیند نه تنها مدل های بهتری تولید کند، بلکه با افزایش اندازه پچ، کارایی محاسباتی را افزایش دهد، زیرا میزبان های بیشتری را می توان با وصله های کمتری شبیه سازی کرد.
5. نتیجه گیری ها
مدلهای پشتیبانی تصمیمگیری محاسباتی برای فعالیتهای نظارت زیستی به انعطافپذیری زیادی در کیفیت و انواع دادهها نیاز دارند که باید برای ارائه نتایج عملی برای کنترل شیوع بیماریهای عفونی ترکیب شوند. ما یک چارچوب مدلسازی ریاضی ارائه کردهایم که گسترش بیماریهای عفونی را در میان گونههای متعدد میزبان توصیف میکند، در حالی که ناهمگونی فضایی را در شبکه تماس میزبان، نقشهایی که گونههای مختلف در اپیدمیولوژی بازی میکنند، و اثرات مداخله انسانی را شامل میشود. ما نشان دادهایم که با استفاده از تجمعهای جمعیتی در سطح شهرستان با وفاداری پایین و در دسترس عموم و پارامترهای استخراجشده از ادبیات بیولوژیکی و اپیدمیولوژیک، میتوان دو پاتوژن با علاقه بالا، FMD و HPAI را شبیهسازی کرد و پیامدهای آنها را به طور عادلانه توصیف کرد.
ما همچنین می بینیم که هر یک از جزئیات مختلف گنجانده شده در ساختار مدل اهمیت دارد – وابستگی متفاوت به ویژگی های بیماری، استراتژی های کاهش متفاوت، گونه های میزبان مختلف، ناهمگونی جغرافیایی در توزیع جمعیت میزبان – همگی بر نتیجه اپیدمی تأثیر می گذارند. مدلهای بسیار دقیق که انطباق با بیماریها و میزبانهای مختلف دشوار است، ممکن است به دلیل عدم قطعیت در هر یک از عوامل فوق که هر یک میتوانند بر نتیجه تأثیر بگذارند، پاسخهای متفاوتی در انتشار اپیدمیها ارائه دهند. مدل های دقیق تر ممکن است پیش بینی های بهتری را در مقیاس محلی به ویژه ارائه دهند. با این حال، دانستن مناطق حیاتی برای انتشار سریع و گستردهتر این بیماری به دانستن اینکه کجا باید استراتژیهای کاهش و کنترل حرکت حیوانات را اعمال کرد و کجا برای مدلهای دقیقتر تلاش کرد، کمک میکند.
در نتیجه، چارچوب مدل ارائه شده در اینجا را می توان به عنوان ارائه “خط اول دفاع” برای مدیریت و پیشگیری از شیوع بیماری های عفونی در حال ظهور در دام و طیور در ایالات متحده در نظر گرفت. ماهیت کلی معماری مدل را می توان با گونه های میزبان متعدد و پاتوژن های متعدد تطبیق داد در حالی که جنبه های مهم ناهمگونی فضایی و حمل و نقل از راه دور را در بر می گیرد. این پارامتر با داده های عمومی در دسترس است و راهبردهای کاهش متداول را در خود جای داده است. علاوه بر این، این مدل از نظر محاسباتی کارآمد است، بنابراین میتواند به راحتی چندین بار برای سناریوهای مختلف اجرا شود تا بازخورد بلادرنگ را به سیاستگذاران و دانشمندان ارائه دهد. چارچوبهای مدلی مانند این میتواند برای اطلاع رسانی اقدامات کاهشی و نظارتی مورد استفاده قرار گیرد.
ساختار مدل فعلی فقط در مورد بیماری های مستقیم قابل انتقال است. ما قصد داریم مدل را برای ترکیب بیماریهای منتقله از طریق ناقل که توسط کنهها، پشهها، میگها و غیره منتقل میشوند، تطبیق دهیم. تنوع بالا در گونههای ناقل و پراکنشها همراه با وابستگی بسیار نزدیک، اما نه اغلب ساده، آنها به آب و هوا و آب و هوای محلی، افزودهای را به همراه خواهد داشت. چالشهای انطباق این چارچوب مدل ترکیب فصلی و تأثیر آن بر انتقال پاتوژن و رفتار حیوان یا انسان نیز جالب خواهد بود.
منابع
- وولف، ND; دوناوان، CP; Diamond, J. ریشه های بیماری های عفونی اصلی انسان. طبیعت 2007 ، 447 ، 279-283. [ Google Scholar ] [ CrossRef ]
- Kao، RR نقش مدلسازی ریاضی در کنترل اپیدمی FMD در سال 2001 در بریتانیا. Trends Microbiol. 2002 ، 10 ، 279-286. [ Google Scholar ] [ CrossRef ]
- Fasina, FO; سردار، م.م. Bisschop، SPR پیامدهای هزینه مالی آنفولانزای فوق حاد پرندگان H5N1 در نیجریه. Onderstepoort J. Vet. Res. 2008 ، 75 ، 39-46. [ Google Scholar ]
- گلوبیگ، آ. استاوباخ، سی. بیر، م. کوپن، یو. فیدلر، دبلیو. نیبورگ، ام. ویلکینگ، اچ. استاریک، ای. تیفکه، جی پی; ورنر، او. و همکاران جنبه های اپیدمیولوژیک و پرنده شناسی شیوع ویروس آنفلوانزای فوق حاد پرندگان H5N1 از دودمان آسیایی در پرندگان وحشی در آلمان، 2006 و 2007. Transbound. ظهور. دیس 2009 ، 56 ، 57-72. [ Google Scholar ] [ CrossRef ]
- هشدار و پاسخ به اپیدمی و همه گیر سازمان جهانی بهداشت. در دسترس آنلاین: http://www.who.int/influenza/human_animal_interface/H5N1_cumulative_table_archives/en/index.html (در 30 دسامبر 2012 قابل دسترسی است).
- شینده، وی. پل ها، CB; Uyeki، TM; شو، بی. بالیش، ا. خو، X. لیندستروم، اس. گوباروا، LV; ديده، وي. Garten، RJ; و همکاران آنفولانزای خوکی سهگانهای (H1) در انسان در ایالات متحده، 2005-2009. N. Engl. جی. مد. 2009 . [ Google Scholar ] [ CrossRef ]
- لیپسیچ، ام. برگستروم، CT تفسیر دعوت شده: ردیابی در زمان واقعی اقدامات کنترلی برای عفونت های در حال ظهور. صبح. J. Epidemiol. 2004 ، 160 ، 517-519. [ Google Scholar ] [ CrossRef ]
- Cowling، BJ; هو، ال ام. Leung، GM اثربخشی اقدامات کنترلی در طول اپیدمی SARS در پکن: مقایسه منحنی R t و منحنی اپیدمی. اپیدمیول. آلوده کردن 2008 ، 136 ، 562-566. [ Google Scholar ]
- فریزر، سی. دانلی، کالیفرنیا؛ کوچمز، اس. Hanage، WP; ون کرخوف، MD؛ Hollingsworth، TD; گریفین، جی. باگالی، RF; جنکینز، HE; لیون، ای جی; و همکاران پتانسیل همه گیری یک سویه آنفولانزای a (H1N1): یافته های اولیه. Science 2009 ، 324 ، 1557-1561. [ Google Scholar ] [ CrossRef ]
- گارسکه، تی. لگراند، جی. دانلی، کالیفرنیا؛ وارد، اچ. کوچمز، اس. فریزر، سی. فرگوسن، NM; غنی، AC ارزیابی شدت همه گیری جدید آنفلوانزا a/H1N1. برادر پزشکی J. 2009 , 339 . [ Google Scholar ] [ CrossRef ]
- بتنکورت، LMA؛ ریبیرو، آر.ام. چاول، جی. لانت، تی. Castillo-Chavez, C. Towards Real Time Epidemiology: Assimilation Data Assimilation, Modeling and Anomaly Detection of Health Surveillance Data Streams. در مجموعه مقالات اطلاعات اطلاعاتی و امنیتی 2007: نظارت زیستی. دومین کارگاه NSF، نیوبرانزویک، نیوجرسی، ایالات متحده آمریکا، 22 مه 2007; صص 79-90.
- واتس، دی جی؛ استروگاتز، SH پویایی جمعی شبکه های “جهان کوچک”. طبیعت 1998 ، 394 ، 440-442. [ Google Scholar ] [ CrossRef ]
- کرماک، WO; McKendrick، AG سهمی در نظریه ریاضی اپیدمی ها. I. Proc. R. Soc. لندن. یک 1927 . [ Google Scholar ] [ CrossRef ]
- فرگوسن، NM; دانلی، کالیفرنیا؛ اندرسون، RM اپیدمی تب برفکی در بریتانیای کبیر: الگوی گسترش و تأثیر مداخلات. Science 2001 , 292 , 1155-1160. [ Google Scholar ] [ CrossRef ]
- گائو، دی. کوسنر، سی. Cantrell، RS; Beier, JC; Ruan, S. مدل سازی گسترش فضایی تب دره شکاف در مصر. گاو نر ریاضی. Biol. 2013 ، 75 ، 523-542. [ Google Scholar ] [ CrossRef ]
- گیلبرت، ام. آکتاس، س. محمد، ح. رودر، پی. سامپشن، ک. توفان، م. Slingenbergh, J. الگوهای گسترش و تداوم بیماری تب برفکی انواع A، O و Asia-1 در ترکیه: یک رویکرد فرا جمعیتی. اپیدمیول. آلوده کردن 2005 ، 133 ، 537-545. [ Google Scholar ] [ CrossRef ]
- تران، سی. یوست، آر. یاناگیدا، جی. ساکسنا، س. فاکس، جی. Sultana، N. مدلسازی وقوع فضایی-زمانی آنفلوانزای فوقالعاده پرندگان H5N1: مطالعه موردی در دلتای رودخانه قرمز، ویتنام. ISPRS Int. J. Geo-Inf. 2013 ، 2 ، 1106-1121. [ Google Scholar ] [ CrossRef ]
- آرینو، جی. جردن، آر. ون دن دریش، P. قرنطینه در یک مدل اپیدمی چند گونه با پویایی فضایی. ریاضی. Biosci. 2007 ، 206 ، 46-60. [ Google Scholar ] [ CrossRef ]
- نیو، تی. گاف، HD; Papelis، YE; هارتلی، DM یک مدل اپیدمیولوژیک تب دره شکاف با پویایی فضایی. محاسبه کنید. ریاضی. روش ها Med. 2012 . [ Google Scholar ] [ CrossRef ]
- شو، ال. اسکات، اچ ام. Cohnstaedt، LW; Scoglio, C. یک رویکرد متا جمعیت مبتنی بر شبکه برای مدلسازی اپیدمیهای تب دره شکاف. جی. تئور. Biol. 2012 ، 306 ، 129-144. [ Google Scholar ]
- بلشام، GJ ویژگی های متمایز ویروس تب برفکی، عضوی از خانواده پیکورناویروس. جنبه های سنتز پروتئین ویروس، پردازش پروتئین و ساختار. برنامه بیوفیز. مول. Biol. 1993 ، 60 ، 241-260. [ Google Scholar ]
- ریواس، آل. کونزبرگ، بی. چاول، جی. اسمیت، SD; هیمن، جی.ام. شواگر، SJ پراکندگی اپیدمی بیماری تب برفکی با واسطه انسان: خوشه های بیماری و ناقل. J. Vet. پزشکی سری B 2006 ، 53 ، 1-10. [ Google Scholar ]
- کیلینگ، ام. Woolhouse، MEJ; شاو، دی جی؛ متیوز، ال. چیس تاپینگ، م. هایدون، دی.تی. کرنل، اس جی; کپی، جی. ویلسمیت، جی. گرنفل، BT Dynamics 2001 اپیدمی تب و دهان انگلستان: پراکندگی تصادفی در یک چشم انداز ناهمگن. Science 2001 , 294 , 813-817. [ Google Scholar ] [ CrossRef ]
- سرویس ملی آمار کشاورزی NASS. در دسترس آنلاین: http://www.nass.usda.gov/Census (در 2 سپتامبر 2009 قابل دسترسی است).
- مانور، سی. مک ماهون، بی. فیر، جی. هیمن، جی. براون، م. LaBute، M. ویژگیهای بیماری، جغرافیا، و استراتژیهای کاهش در گسترش شبیهسازی آفت گاو در سراسر ایالات متحده. دامپزشک Res. 2011 ، 42 . [ Google Scholar ] [ CrossRef ]
- مایر، RF؛ Knudsen، RC بیماری پا و دهان: مروری بر ویروس و علائم. جی. محیط زیست. سلامت 2001 ، 64 ، 21-23. [ Google Scholar ]
- الکساندرسن، اس. دونالدسون، هوش مصنوعی مطالعات بیشتری برای تعیین کمیت دوز آئروسلهای طبیعی ویروس تب برفکی برای خوکها انجام شد. اپیدمیول. آلوده کردن 2002 ، 128 ، 313-323. [ Google Scholar ]
- کالنز، ام. د کلرک، ک. گرویا، م. Danes، M. تشخیص بیماری تب برفکی با واکنش زنجیره ای پلیمراز رونویسی معکوس و جداسازی ویروس در گوسفندان تماسی بدون علائم بالینی بیماری تب برفکی. دامپزشک Q. 1998 , 20 , S37–S40. [ Google Scholar ] [ CrossRef ]
- بارنت، PV; Cox, SJ نقش نشخوارکنندگان کوچک در اپیدمیولوژی و انتقال بیماری تب برفکی. دامپزشک J. 1999 ، 158 ، 6-13. [ Google Scholar ] [ CrossRef ]
- هیوز، جی جی; میولت، وی. هایدون، دی.تی. کیچینگ، RP; دونالدسون، هوش مصنوعی؛ Woolhouse، ME عبور سریالی ویروس تب برفکی در گوسفند نشان دهنده کاهش سطوح ویرمی در طول زمان است. جی ژنرال ویرول. 2002 ، 83 ، 1907-1914. [ Google Scholar ]
- ناولز، نیوجرسی؛ ساموئل، AR اپیدمیولوژی مولکولی ویروس تب برفکی. ویروس Res. 2003 ، 91 ، 65-80. [ Google Scholar ] [ CrossRef ]
- دونالدسون، هوش مصنوعی؛ الکساندرسن، اس. پیش بینی گسترش بیماری تب برفکی توسط ویروس هوابرد. Revue Sci. فنی خاموش بین المللی اپیزوت. 2002 ، 21 ، 569-575. [ Google Scholar ]
- الکساندرسن، اس. کوان، ام. مورفی، سی. نایت، جی. Zhang, Z. مطالعات پارامترهای کمی دفع و انتقال ویروس در خوکها و گاوهای آلوده تجربی به ویروس تب برفکی. J. Comp. پاتول. 2003 ، 129 ، 268-282. [ Google Scholar ] [ CrossRef ]
- فروشندگان، RF; Parker, J. دفع ویروس تب برفکی از طریق هوا. جی. هیگ. 1969 ، 67 ، 671. [ Google Scholar ] [ CrossRef ]
- Shields, ME Agro-terorism, Biotechnology, and Biosis. جی. آگریک. Food Inf. 2003 ، 5 ، 19-23. [ Google Scholar ] [ CrossRef ]
- کاریلو، سی. لو، ز. Borca، MV; واگنوزی، آ. کوتیش، جی اف. Rock، DL تنوع ژنتیکی و فنوتیپی ویروس تب برفکی در طول گذرهای سریال در یک میزبان طبیعی. جی. ویرول. 2007 ، 81 ، 11341-11351. [ Google Scholar ] [ CrossRef ]
- اورسل، ک. بوما، ا. دکر، ا. Stegeman، JA; de Jong، MCM انتقال ویروس بیماری تب برفکی در طول دوره نهفتگی بیماری در خوکها، برهها، گوسالهها و گاوهای شیری. قبلی دامپزشک پزشکی 2009 ، 88 ، 158-163. [ Google Scholar ] [ CrossRef ]
- گلد، WT؛ Nfon، CK؛ توکا، FN فرار ایمنی در طی عفونت ویروسی بیماری تب برفکی خوکی. ایمونول. Rev. 2008 , 225 , 85-95. [ Google Scholar ] [ CrossRef ]
- کیچینگ، RP; هیوز، GJ تغییرات بالینی در بیماری پا و دهان: گوسفند و بز. Revue Sci. فنی خاموش بین المللی اپیزوت. 2002 ، 21 ، 505-512. [ Google Scholar ]
- دانلی، کالیفرنیا؛ اندرسون، شدت انتقال RM و تأثیر سیاستهای کنترل بر اپیدمی تب و دهان در بریتانیای کبیر. طبیعت 2001 ، 413 ، 542-548. [ Google Scholar ] [ CrossRef ]
- کوان، ام. مورفی، سی ام. ژانگ، ز. الکساندرسن، S. عوامل تعیین کننده پویایی ویروس تب برفکی اولیه در خوک ها. J. Comp. پاتول. 2004 ، 131 ، 294-307. [ Google Scholar ]
- الکساندرسن، اس. ژانگ، ز. دونالدسون، هوش مصنوعی جنبههای تداوم ویروس تب برفکی در حیوانات: مشکل ناقل. میکروب ها عفونی می کنند. 2002 ، 4 ، 1099-1110. [ Google Scholar ] [ CrossRef ]
- ونگ، اس. کوغلان، بی. مردی، س. هال، دی. سنگ، اچ. لی، اس. میلر، ام جی; بوچی، پی. فروهلیچ، ی. Dufourcq، JB; و همکاران فرکانس پایین انتقال ویروس H5Ni از طیور به انسان، کامبوج جنوبی، 2005. Emerg. آلوده کردن دیس 2006 ، 12 ، 1542-1547. [ Google Scholar ] [ CrossRef ]
- مارین، سی. تشخیص Lainez، M. Salmonella در مدفوع در طی پرورش جوجه های گوشتی و پس از انتقال زنده به کشتارگاه. مرغ. علمی 2009 ، 88 ، 1999-2005. [ Google Scholar ] [ CrossRef ]
- ماس، م. ایمادا، تی. ناکامورا، ک. تانیمورا، ن. ایمای، ک. سوکاموتو، ک. یاماگوچی، اس. ارزیابی تجربی بیماری زایی ویروس آنفولانزای H5N1 جدا شده در ژاپن. پرندگان دیس. 2005 ، 49 ، 582-584. [ Google Scholar ] [ CrossRef ]
- شورتریج، KF; ژو، NN; گوان، ی. گائو، پی. ایتو، تی. کاوائوکا، ی. کودیهالی، س. کراوس، اس. مارکول، دی. مورتی، KG; و همکاران مشخصات ویروس آنفلوانزای پرندگان H5N1 از طیور در هنگ کنگ. ویروس شناسی 1998 ، 252 ، 331-342. [ Google Scholar ] [ CrossRef ]
- اوسترهاوس، آ. لهستان، واکسن های GA علیه آنفولانزای فصلی و پرندگان: پیشرفت های اخیر. واکسن 2008 ، 26 ، D1-D2. [ Google Scholar ]
- بوما، ا. کلاسن، آی. ناطیه، ک. کلینکنبرگ، دی. دانلی، کالیفرنیا؛ کوچ، جی. van Boven, M. برآورد پارامترهای انتقال ویروس آنفلوانزای مرغی H5N1 در جوجه ها. PLoS Pathog. 2009 . [ Google Scholar ] [ CrossRef ]
- Latorre-Margalef، N. گونارسون، جی. مونستر، وی جی. Fouchier، RAM; اوسترهاوس، ا. المبرگ، جی. اولسن، بی. والنستن، ا. Haemig، PD; فرانسون، تی. و همکاران اثرات عفونت ویروسی آنفولانزا بر اردک های اردک اردک مهاجر. Proc. R. Soc. B-Biol. علمی 2009 ، 276 ، 1029-1036. [ Google Scholar ] [ CrossRef ]
- ساساکی، تی. کوکومای، ن. اوهگیتانی، ت. ساکاموتو، آر. تاکیکاوا، ن. لین، ZF؛ اوکاماتسو، ام. ساکودا، ی. Kida، H. ایمنی طولانی مدت در جوجه ها ناشی از یک شات واکسن آنفولانزا تهیه شده از ذرات غیر فعال ویروس H5N1 غیر بیماری زا در برابر چالش با ویروس آنفولانزای پرندگان بسیار بیماری زا. واکسن 2009 ، 27 ، 5174-5177. [ Google Scholar ] [ CrossRef ]
- کگل، سی. واسیلنکو، جی. آدامز، SC; کاردونا، سی جی; تان لانگ، تی. تونگ، ن. اسپکمن، ای. سوارز، دی.ال. اسمیت، دی. شپرد، ای. و همکاران تفاوت در بیماری زایی، پاسخ به واکسیناسیون، و پاسخ های ایمنی ذاتی در انواع مختلف اردک های آلوده به ویروس خطرناک H5N1 آنفولانزای پرندگان بسیار بیماری زا از ویتنام. پرندگان دیس. 2012 ، 56 ، 479-487. [ Google Scholar ]
- نیمکت، RB; پاتل، اس ام. Wade-Bowers، CL; نینو، دی. PLoS One 2012 . [ Google Scholar ] [ CrossRef ]
- Leung، YHC; لوک، جی. سیا، س.-ف. وو، Y.-O. هو، سی.-ک. چاو، K.-C.; تانگ، اس.-سی. گوان، ی. Peiris، JSM چالش آزمایشی مرغ واکسینه شده با واکسن های H5 تجاری موجود، از دست دادن حفاظت را برای برخی از سویه های H5N1 آنفولانزای پرندگان بسیار بیماری زا که در هنگ کنگ/چین در گردش هستند، نشان می دهد. واکسن 2013 ، 31 ، 3536-3542. [ Google Scholar ] [ CrossRef ]
- ایوامی، س. تاکوچی، ی. لیو، XN; ناکائوکا، S. گسترش جغرافیایی مقاومت به واکسن در اپیدمی های آنفلوانزای مرغی. جی. تئور. Biol. 2009 ، 259 ، 219-228. [ Google Scholar ] [ CrossRef ]
- لبربنشون، سی. آلبسپی، اف. Brochet, A.-L.; مادربزرگ، V. رنو، اف. فریتز، اچ. گرین، ای جی؛ توماس، اف. ون در ورف، اس. اوبری، پی. و همکاران انتشار ویروس آنفولانزای پرندگان توسط گیاه معمولی ( anas crecca ) در اروپا. PLoS One 2009 , 4 . [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- ریواس، آل. چاول، جی. شواگر، اس جی; Fasina, FO; Hoogesteijn، AL; اسمیت، SD; Bisschop، SPR; اندرسون، KL; Hyman, JM Lessons from Nigeria: نقش جادهها در پیشرفت جغرافیایی-زمانی ویروس آنفلوانزای مرغی (H5N1). اپیدمیول. آلوده کردن 2010 ، 138 ، 192-198. [ Google Scholar ] [ CrossRef ]
- جنبش های دام بین ایالتی USDA. در دسترس آنلاین: http://www.ers.usda.gov/Data/InterstateLivestockMovements/ (در 4 مارس 2009 قابل دسترسی است).
- برنهام، ام آر. پیبلز، ED; برانتون، اس ال. جونز، ام اس؛ PD، G. اثرات تلقیح مایکوپلاسما گالی سپتیکوم با سویه f در سن دوازده هفتگی بر خصوصیات خونی مرغ های تخمگذار تجاری. مرغ. علمی 2003 ، 82 ، 1397-1402. [ Google Scholar ] [ CrossRef ]
- بیتس، TW; نجار، TE; Thurmond، MC تجزیه و تحلیل سود-هزینه واکسیناسیون و کشتار پیشگیرانه به عنوان وسیله ای برای ریشه کنی بیماری تب برفکی. صبح. J. Vet. Res. 2003 ، 64 ، 805-812. [ Google Scholar ] [ CrossRef ]
- Doeschl-Wilson، AB; دیویدسون، آر. کانینگتون، جی. رافسج، تی. Hutchings، MR; Villanueva, B. مفاهیم تنوع ژنتیکی میزبان بر خطر و شیوع بیماری های عفونی منتقل شده از طریق محیط. ژنتیک 2011 ، 188 ، 683-693. [ Google Scholar ] [ CrossRef ]
- Rasmussen, S. مدلسازی تنوع فضایی گسسته در اپیدمیولوژی با SAS با استفاده از Glimmix. محاسبه کنید. Methods Progr. بیومد. 2004 ، 76 ، 83-89. [ Google Scholar ] [ CrossRef ]
- مارتین، وی. شوالیه، وی. سکاتو، پ. آنیامبا، ا. دی سیمون، ال. لوبروت، جی. د لا روک، اس. دومنک، جی. تأثیر تغییرات آب و هوا بر اپیدمیولوژی و کنترل تب دره ریفت. Revue Sci. فنی – خاموش. بین المللی اپیزوت. 2008 ، 27 ، 413-426. [ Google Scholar ]
- هاگرمن، AD; بخش، نماینده مجلس؛ اندرسون، DP; لونی، جی سی؛ مک کارل، BA ارزش قابلیت ردیابی موثر سریع: مطالعه موردی بیماری پا و دهان در دشتهای تگزاس. قبلی دامپزشک پزشکی 2013 ، 110 ، 323-328. [ Google Scholar ] [ CrossRef ]
- الباکیدزه، ال. هایفیلد، ال. وارد، م. مک کارل، کارشناسی; Norby، B. تجزیه و تحلیل اقتصادی استراتژی های کاهش برای معرفی FMD در مناطق بسیار متمرکز تغذیه حیوانات. کشیش کشاورزی اقتصاد 2009 ، 31 ، 931-950. [ Google Scholar ] [ CrossRef ]
- بتنکورت، LMA؛ Ribeiro، RM تخمین بیزی در زمان واقعی از پتانسیل اپیدمی بیماری های عفونی در حال ظهور. PLoS One 2008 , 3 . [ Google Scholar ] [ CrossRef ]
© 2014 توسط نویسندگان; دارنده مجوز MDPI، بازل، سوئیس. این مقاله یک مقاله با دسترسی آزاد است که تحت شرایط و ضوابط مجوز Creative Commons Attribution (http://creativecommons.org/licenses/by/3.0/) توزیع شده است.
بدون نظر