

خلاصه
بر اساس دستورالعمل اتحادیه اروپا برای زیستگاه ها، گرمسیری ها “زیستگاه های طبیعی مورد علاقه جامعه” هستند. هدف مدیریت Heathland حفاظت از این زیستگاه های تهدید شده توسط تغییرات مختلف، از جمله فرآیندهای متوالی منجر به پوشش گیاهی جنگل است. ما پویایی گونههای چوبی را به ضرر گرمک طی یک دوره 60 ساله در جنگل فونتنبلو بررسی میکنیم و اثرات انواع خاک، عمق خاک و پارامترهای توپوگرافی را بر پایداری گرمک بررسی میکنیم. ما تغییرات پوشش جنگلی را بین سالهای 1946 و 2003 با مقایسه نقشههای پوشش گیاهی به دست آمده از عکسهای هوایی همراه با تجزیه و تحلیل GIS ارزیابی میکنیم. نتایج نشان میدهد که بیش از 75 درصد از هتکها در طول سالهای 1946-2003 به دلیل استعمار درختان هولدر متروکه از بین رفته است. ما تفاوتهایی را در پویایی استعمار بین درختان مخروطی و برگریز شناسایی کردیم. استعمار هیتلند توسط گونه های مخروطی در 20 سال گذشته دوره مطالعه ما سریعتر بود. تجاوز درختان در نواحی رو به شمال و در نواحی لوویزول های اسیدی سریعتر بود. در حالی که این دینامیک در خاکهای ماسهسنگ اسیدی بسیار آهسته بود، پایداری هلند در خاکهای کمعمق در مناطق هموار و رو به جنوب اهمیت بیشتری داشت. مطالعه ما این پتانسیل را دارد که به مدیران زمین در انتخاب مناطق گرمسیری کمک کند که با تمرکز بر مناطق و شرایط فضایی که از استعمار جنگل ها جلوگیری می کند و از این رو به پایداری بلندمدت گرمسیری کمک می کند، حفاظت و/یا بازسازی آنها آسان تر باشد. در حالی که این دینامیک در خاکهای ماسهسنگ اسیدی بسیار آهسته بود، پایداری هلند در خاکهای کمعمق در مناطق هموار و رو به جنوب اهمیت بیشتری داشت. مطالعه ما این پتانسیل را دارد که به مدیران زمین در انتخاب مناطق گرمسیری کمک کند که با تمرکز بر مناطق و شرایط فضایی که از استعمار جنگل ها جلوگیری می کند و از این رو به پایداری بلندمدت گرمسیری کمک می کند، حفاظت و/یا بازسازی آنها آسان تر باشد. در حالی که این دینامیک در خاکهای ماسهسنگ اسیدی بسیار آهسته بود، پایداری هلند در خاکهای کمعمق در مناطق هموار و رو به جنوب اهمیت بیشتری داشت. مطالعه ما این پتانسیل را دارد که به مدیران زمین در انتخاب مناطق گرمسیری کمک کند که با تمرکز بر مناطق و شرایط فضایی که از استعمار جنگل ها جلوگیری می کند و از این رو به پایداری بلندمدت گرمسیری کمک می کند، حفاظت و/یا بازسازی آنها آسان تر باشد.
کلید واژه ها:
GIS ; تغییر پوشش زمین ; حفاظت از تنوع زیستی ; منطقه حفاظت شده ؛ جانشینی ثانویه ; هیتلند
1. معرفی
تغییرات جهانی اکوسیستم های طبیعی را تهدید می کند که در حال فروپاشی و حتی به طور کامل ناپدید می شوند [ 1 ]. حفاظت از آنها یک اولویت به منظور توقف از دست دادن تنوع زیستی [ 2 ] است و در حال حاضر توسط کنوانسیون ها و برنامه هایی که هدف آنها حفظ و احیای زیستگاه های طبیعی است، تضمین شده است. روشهای مدیریتی مختلف باید برای حفظ زیستگاههای در معرض تهدید در وضعیت حفاظتی مطلوب [ 3 ]، مانند کنترل جانشینی طبیعی و ساختار فیزیکی پوشش گیاهی، برای حفظ گونهها یا مجموعههای گونهای مورد توجه حفاظت استفاده شود [ 4 ]. زیستگاه گرمک اروپایی نمونه ای معمولی از زیستگاهی است که در آن چنین مدیریت فعالی مورد نیاز است. مناطق گرمسیری اروپا تحت سلطه گونه های گیاهی مشخصی هستند،Calluna vulgaris (L.) Hull ، [ 5 ]، که منبع اصلی برای گونه های تخصصی پرندگان و بی مهرگان است [ 6 ، 7 ]. این زیستگاه محدود به خاکهای اسیدی و فقیر از مواد مغذی [ 8 ] است و یک مرحله پیشگام در توالی طبیعی است [ 9 ]] به جز مواقعی که در برخی از نواحی ساحلی به طور طبیعی تالاب ها رخ می دهند. در طول 3000 سال گذشته، مناطق وسیعی از مناطق گرمسیری اروپا در سرتاسر شمال غربی اروپا توسط شیوههای سنتی کشاورزی – شبانی حفظ شدهاند. از آنجا که تشدید کشاورزی در دهه 1950 رخ داد، این کاربری های سنتی تقریباً به طور کامل ناپدید شدند. در نتیجه، مناطق گرمسیری به شدت کاهش یافته است زیرا پوشش گیاهی در خاک های اسیدی و فقیر از مواد مغذی توسط گونه های چوبی پوشیده شده است. از آنجایی که در ضمیمه I دستورالعمل زیستگاه های اتحادیه اروپا در سال 1992 به عنوان یک نوع زیستگاه طبیعی مورد علاقه جامعه نامگذاری شد، برنامه های مدیریت زیستگاه به طور فزاینده ای با هدف حفظ گونه ها و زیستگاه های گرمسیری [ 10 ] انجام می شود.] و نگهداری آنها در وضعیت مطلوب حفاظتی. با این وجود، بسیاری از مطالعات نشان میدهد که گسترش جنگلها در گرمابه علیرغم اعمال اقدامات مختلف برای حفظ این زیستگاه، مانند سوزاندن تجویز شده و قلمههای مکانیکی [ 11 ]. تحت شرایط غالب تغییرات آب و هوایی و رسوب نیتروژن، حفاظت از گرمسیری دشوارتر میشود و به شیوههای مدیریتی فشردهتر [ 12 ] با افزایش هزینه نیاز دارد ، زیرا خشکسالی و تغییرات بارش فصلی بر تعادل رقابتی بین گونهها تأثیر میگذارد. از سوی دیگر، افزایش میزان رسوب نیتروژن در اکوسیستمهای زمینی، حاصلخیزی خاک و توانایی رقابتی علفها را بهبود میبخشد، اما به ضرر زمینهای گرمسیری است.14 ، 15 ].
در این زمینه، شناسایی تأثیر تغییرپذیری فضایی در خاک و توپوگرافی بر پویایی موزائیک کوههای جنگلی به منظور تعیین مناسبترین روشهای مدیریتی برای حفاظت طولانیمدت این موزاییک یک ضرورت واقعی میشود. پویایی جوامع گیاهی به طور مستقیم با تنوع نوع خاک مرتبط است، زیرا منابع معدنی خاک یک عامل ساختاری برای سازماندهی پوشش گیاهی است [ 16 ]. در واقع، تنوع فضایی در عوامل فیزیوگرافی میتواند پوشش گیاهی [ 17 ، 18 ، 19 ] و جمعیتهای گیاهی [ 20 ] را نیز کنترل کند، که میتواند بر پویایی جنگل به ضرر گرمکها تأثیر بگذارد.
در جنگل فونتنبلو در 50 کیلومتری جنوب پاریس، فرانسه، مناطق گرمسیری هنوز به صورت تکههای کوچکی که در مناطق جنگلیتر جاسازی شدهاند، وجود دارد و یک منظره موزاییکی بسیار پیچیده را تشکیل میدهد که میزبان گونههای خاصی است. در این جنگل، وجود خاکهای شنی اسیدی و کشاورزی مرسوم، باعث ایجاد و نگهداری از گرما در طول هزاران سال شده است. رها شدن کاربریهای اراضی اجدادی از نیمه دوم قرن بیستم باعث کاهش گرماگردی شده است. در حال حاضر، 1400 هکتار از هتک باقی مانده است، در تکه های تکه تکه شده در جنگل بلوط کاج که در یک ذخیره گاه زیستی مدیریت شده قرار گرفته است. از اوایل دهه نود، اداره ملی جنگل (ONF)، به دنبال تعهدات فرانسه در رابطه با دستورالعمل های اتحادیه اروپا، تلاش کرده است تا با قطع منظم گیاهان جدید گونه های چوبی، بسیاری از لکه ها را حفظ کند. این تعمیر و نگهداری به دلیل پیکربندی فضایی قطعات گرمسیری که در ماتریس بزرگی از جنگلهای مخروطی و برگریز پراکنده شدهاند، روز به روز دشوارتر میشود. اقتصادیترین روشهای مدیریتی، مانند قلمه زدن منظم مکانیکی و حذف گونههای چوبی، برای حفظ گونههای خاص در یک قطعه گرمسیری در وسط یک ماتریس جنگل، همانطور که در مطالعات قبلی نشان دادیم، کافی نیستند.11 ، 21 ]. این شرایط نیازمند روش های اضافی برای حفظ بهتر این زیستگاه است.
در مطالعه حاضر، ما از تجزیه و تحلیل مکانی-زمانی مبتنی بر عکسهای هوایی برای توصیف تغییرات در موزاییک جنگلها در طی یک دوره 60 ساله استفاده میکنیم تا فرآیندهای استعمار مجدد درختان جنگلی را در هولاد متروکه درک کنیم. در این مطالعه همچنین به دنبال بررسی تأثیر ویژگیهای خاک (نوع و عمق خاک) و عوامل فیزیوگرافیک (شیب و جنبه) بر تغییرات پوشش زمین در این منطقه بودیم.
2. مواد و روشها
2.1. سایت مطالعه
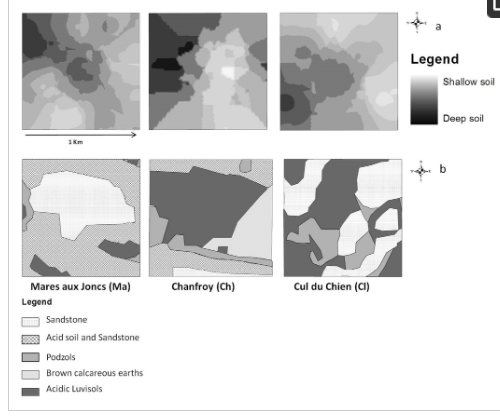
جنگل دولتی به نام “Trois Pignons” با 3307 هکتار، بخشی از جنگل فونتنبلو است ( شکل 1 ) و از موزاییکی از گرمسیری و جنگل تشکیل شده است: 83 هکتار از هتک های مدیریت شده و تقریباً 540 هکتار از گرمک ها با کمتر. بیش از 10٪ پوشش چوبی در ماتریسی از جنگل های مخروطی و برگریز قرار دارد. ما سه سایت میدانی به مساحت 1 کیلومتر مربع را انتخاب کردیممشخصه هر کدام، استعمار مجدد مناطق گرمسیری توسط جنگل است. هر سایت در مرحله متفاوتی از پویایی هالند-جنگل قرار دارد، زیرا میدانیم که هلند در سال 1946 در این سه مکان وجود داشته است. سایت ها نیز از نظر شرایط اکولوژیکی متفاوت هستند. اولین سایت Mares aux Joncs (“Ma”) است که یک چشم انداز Stampian با رخنمون های ماسه سنگ است که محیط های هیدرومورفیک بیش از حد اسیدی را ایجاد می کند. سایت دوم، Chanfroy (“Ch”)، یک دشت کم ارتفاع با مخلوطی از ماسه های سیلیسی و شن های آهکی است، در حالی که سایت 3 Cul du Chien (“Cl”) است، یکی دیگر از منظره های Stampian متشکل از ماسه های سیلیس خالص و اسیدی بیش از حد. .
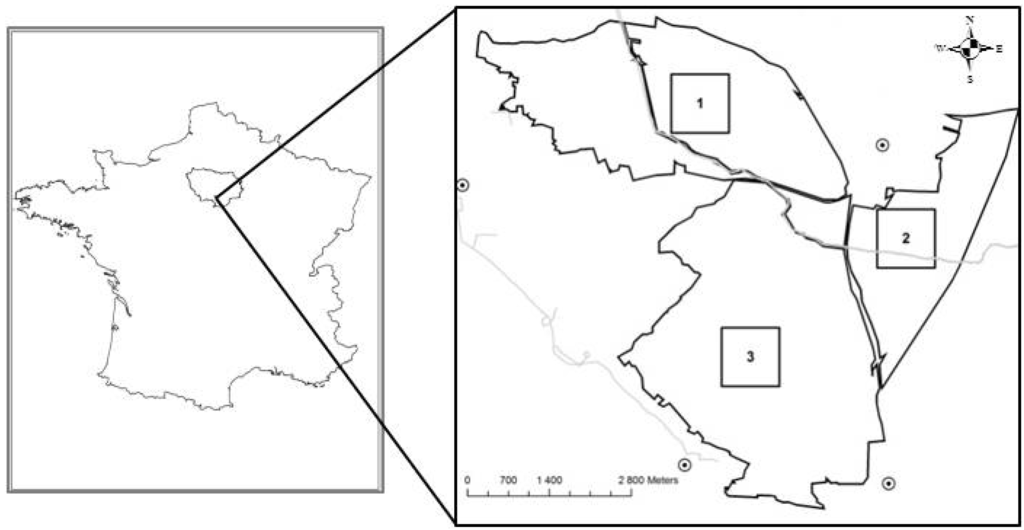
شکل 1. محل سه مکان مطالعه در جنگل (Trois Pignons): 1. Mares aux Joncs (Ma)، 2. Chanfroy (Ch) و 3. Cul du Chien (Cl). خطوط خاکستری نشان دهنده رودخانه ها و نقاط تاریک نشان دهنده شهرها هستند.
2.2. پردازش تصویر
2.2.1. طبقه بندی تغییرات در پوشش گیاهی
استعمار طبیعی مناطق باز با تفسیر عکس از عکسهای هوایی که طی چهار بررسی عکسبرداری هوایی توسط موسسه جغرافیایی ملی در سالهای 1946، 1965، 1985 (عکسهای اصلاحشده سیاه و سفید)، و یک عکس رنگی دیجیتال در سال 2003 مورد مطالعه قرار گرفت. با وضوح مکانی 50 سانتی متر. طبقه بندی پوشش گیاهی بر اساس روش تفسیر بصری بود. هر تصویر با یک شبکه 50 × 50 متر پوشانده شد و سپس هر سلول به یکی از شش کلاس پوشش گیاهی طبقه بندی شد: خاک برهنه، Heathland، علفزار، جنگل های مخروطی، جنگل های برگریز، و جنگل های مختلط. “کلیدهای تفسیری” مورد استفاده در اینجا عبارتند از: 1. رنگ قابل مشاهده، 2. ارتفاع، 3. بافت، و 4. شکل اجسام [ 22 ]]. بررسی های عکاسی هوایی در سال 2003 از BD ORTHO® IGN 2003 شامل تصاویر رنگی واقعی است که بسیار شبیه به آنچه توسط چشم انسان مشاهده می شود، در اینجا برای تمایز بین انواع مختلف پوشش گیاهی استفاده می شود: مخروطی ها به رنگ سبز تیره تا سبز آبی هستند، درختان برگریز هستند. رنگ سبز روشن تا زرد مایل به سبز، لکههای علفها سبز زرد، گرمکها خاکستری مایل به سبز و خاک برهنه تقریباً ماسه سفید است. عکس های هوایی قدیمی از 1946-1985 تصاویر سیاه و سفید هستند. لحن آنها بر اساس سایه های خاکستری است. ما هنوز میتوانستیم بین اجسام روشن مانند درختان برگریز و علفها و اشیاء تیره مانند درختان مخروطی و درختچههای کوتوله تمایز قائل شویم. مناطقی از رنگ همگن در تصویر مربوط به انواع مختلف پوشش زمین به دلیل وجود سایه ها از هم متمایز شدند. که نشان دهنده عناصر جنگلی بودند (به عنوان مثال، درختان بزرگ منفرد، توده های جنگلی). بافت یک عامل مهم در فرآیند تفسیر بصری است که در اینجا بهویژه برای تمایز بین بافت صاف علفزارهای همگن و بافت درشت منطقه گرمسیری استفاده میشود. شکل تاج درختان به عنوان شاخصی برای تشخیص نوع درخت استفاده شد.به عنوان مثال ، برگریز یا مخروطی. مخروطی ها شکل مخروطی دارند در حالی که درختان برگ ریز دارای تاج های پرزدار یا پرزدار هستند.
تبعیض جنگل ها به گونه غالب در هر پیکسل، با آستانه 75 درصد از پیکسل های تحت پوشش، وابسته بود. هنگامی که گونه های مخروطی یا برگریز بالاتر از این آستانه بودند، سلول به ترتیب به عنوان جنگل های مخروطی یا برگریز طبقه بندی می شد. اگر هیچ یک از این گروه ها به آستانه نمی رسید، سلول به عنوان جنگل های مختلط طبقه بندی می شد. این طبقه بندی با استفاده از طبقه بندی پوشش جنگلی (IFN IF 2006، جدول 1 ) [ 23 ] اصلاح شد.

جدول 1. فهرست دسته بندی های مختلف پوشش گیاهی مورد استفاده در فرآیند تفسیر عکس.
برای ارزیابی دقت نقشه های طبقه بندی شده، نتایج طبقه بندی باید با داده های مرجع جمع آوری شده در زمین مقایسه شود [ 24 ]. در منطقه مورد مطالعه ما، “داده های زمینی” برای عکس های تاریخی قدیمی در دسترس نیست. بنابراین، ما دقت نقشههای طبقهبندی را فقط برای پوشش گیاهی در سال 2003 با مقایسه نقشههای طبقهبندی با دادههای جمعآوریشده روی زمین در 75 نقطه جایی که پوشش گیاهی را در سال 2008 بررسی کردیم، ارزیابی میکنیم (به بخش 2.5 مراجعه کنید).برای جزئیات بیشتر). ما در اینجا در نظر می گیریم که تغییرات بین این پنج سال برای گونه های جنگلی که رشد آهسته و طول عمر طولانی دارند ناچیز است. تغییرات برای تکههای تکهتکهشده هولند در این دوره ناچیز است. علاوه بر این، ONF از اوایل دهه نود سعی در حفظ آنها داشته است.
با گروه بندی همه دسته های جنگلی، الگوهای فضایی جانشینی ثانویه را در سه بازه زمانی به دست آوردیم: 1946-1965، 1965-1985، و 1985-2003. پهنههای غیر جنگلی که در اکثر موارد با اقدامات مدیران حفاظت حفظ شدند، از نظر پویایی پوشش گیاهی بهعنوان پهنههای پایدار در نظر گرفته شدند.
2.2.2. ماتریس های انتقالی
ماتریس های انتقال با جدول بندی متقابل نقشه های متوالی به دست آمدند، و ماتریس های انتقالی را برای دوره های زمانی قابل مقایسه، به ترتیب 19، 20 و 18 سال ارائه کردند. این ماتریس ها پویایی بین انواع مختلف پوشش گیاهی را در طول دوره مورد مطالعه نشان می دهد. این تغییرات زمانی و همچنین ابعاد مکانی را در نظر می گیرد. جداول متقابل با استفاده از IDRISI [ 25 ] انجام شد.
2.3. متغیرهای محیطی فضایی
2.3.1. متغیرهای فیزیوگرافی
ما ویژگیهای فیزیوگرافی ثانویه – شیب و جنبه – را با استفاده از ArcGIS [ 26 ] با یک مدل ارتفاعی رقومی در وضوح فضایی 2.5 متر، که در چارچوب این مطالعه بهعنوان لایه ورودی توسعه یافته است، محاسبه کردیم.
2.3.2. بررسی خاک
برای مطالعه خصوصیات خاک از دو رویکرد استفاده شد:
-
داده های عمق خاک، به دست آمده با نمونه برداری مزرعه. اندازه گیری عمق خاک در 75 نقطه، 25 نقطه در هر سایت انجام شد. یک مطالعه زمین آماری به منظور به دست آوردن نقشه شطرنجی عمق خاک بر اساس نقاط مشاهده انجام شد [ 21 ]. برای انجام این کار، مقادیر را در نقاط مشاهده نشده با استفاده از روش کریجینگ درون یابی کردیم. این روش امکان پیش بینی مقادیر مجهول را از داده های مشاهده شده در مکان های شناخته شده فراهم می کند. کریجینگ از واریوگرام ها برای بیان تغییرات فضایی استفاده می کند و با تخمین توزیع فضایی مقادیر پیش بینی شده، خطاهای پیش بینی را به حداقل می رساند [ 27 ، 28 ]]. به دلیل اثرات لبه، مقادیر تخمینی عمق خاک در لبه های کرت ممکن است در معرض سطوح بالاتری از عدم قطعیت نسبت به سایر نقاط داخل مرزهای کرت باشد. سه طبقه از عمق خاک، مطابق تقریبی با افق های خاک مشاهده شده در این منطقه [ 21 ] متمایز شد: خاک های کم عمق (0-20 سانتی متر)، خاک های با عمق متوسط (21-40 سانتی متر)، و خاک های عمیق (بیش از 40 سانتی متر). ). ما از این نقشه درونیابی به عنوان نقشه عمق خاک همانطور که در شکل 2 الف نشان داده شده است استفاده کردیم.
-
داده های نوع خاک برای شناسایی پراکندگی انواع خاک، از نقشه خاک منطقه [ 29 ] با وضوح فضایی 2.5 متر استفاده کردیم. پنج نوع خاک در منطقه مورد مطالعه یافت میشود: خاکهای آهکی قهوهای، ماسهسنگ، خاک اسیدی و ماسهسنگ، پادزولها و لوویزولهای اسیدی، همانطور که در ( شکل 2 ب) نشان داده شده است.
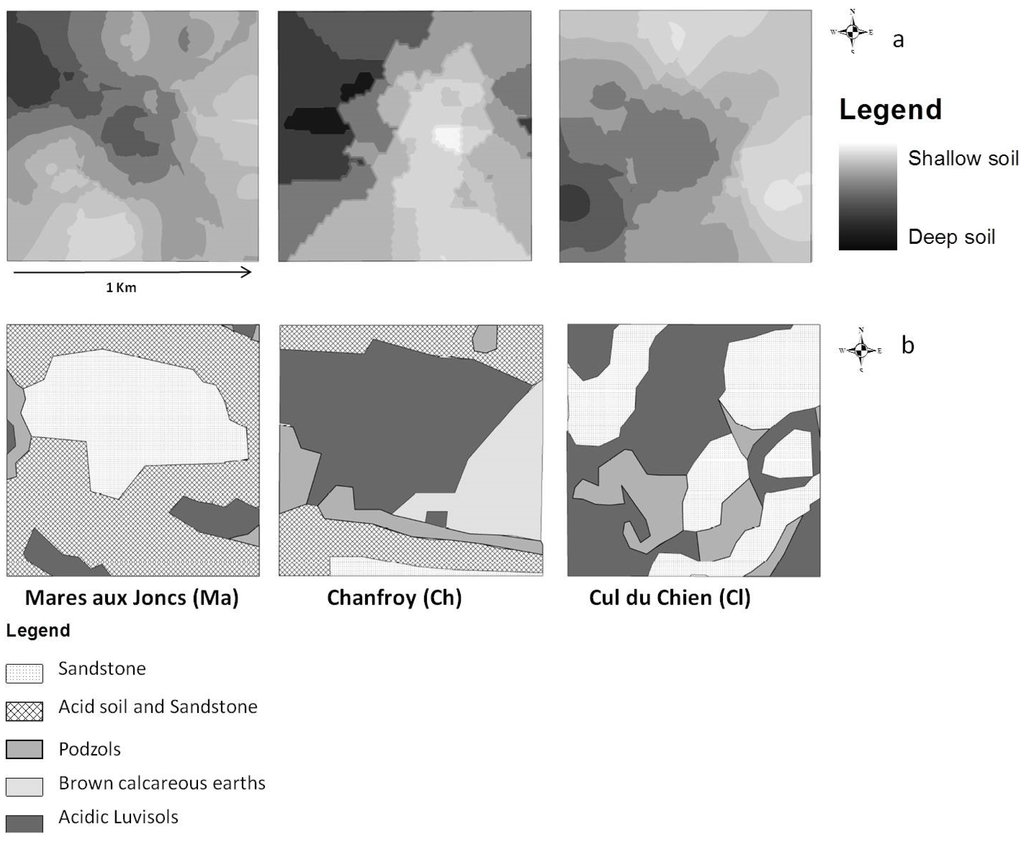
شکل 2. نقشه خاک: ( الف ) عمق خاک، ( ب ) نوع خاک.
2.4. تجزیه و تحلیل داده های مکانی
برای تشخیص تأثیر متغیرهای محیطی فضایی بر گسترش فضایی جانشینی ثانویه، از دو رویکرد آماری استفاده شد. ابتدا، یک ماتریس جدولبندی متقاطع نشاندهنده تعداد پیکسلهایی است که بر اساس دستهبندی بین ترکیبی از دو نقشه شطرنجی طبقهای منطبق هستند. سپس آزمون Chi-square برای ارزیابی اهمیت ارتباط بین نقشههای جانشینی ثانویه و چهار متغیر محیطی فضایی، یعنی عمق خاک، نوع خاک، شیب و جنبه استفاده شد.
پس از آن، با هدف شناسایی شباهتهای مکان بین الگوهای فضایی توالی ثانویه و هر یک از متغیرهای محیطی فضایی، شاخص مکان کاپا (Kloc) را محاسبه کردیم که شاخصی است که توزیع فضایی دستهها را بر روی نقشه نشان میدهد. ، پیشنهاد شده توسط پونتیوس [ 30 ]. برای آماره کاپا (k)، [ 31 ] دستورالعمل هایی را برای تفسیر مقادیر k به شرح زیر ارائه می کند: ضعیف (k < 0)، اندک (0 < k < 0.20)، منصفانه (0.21 < k < 0.40)، متوسط (0.41 < k <0.60)، قابل توجه (0.61< k<0.80)، و تقریباً کامل (0.81 < k <1.00). Kloc در اینجا با استفاده از نرمافزار Map Comparison Kit (MCK) از مؤسسه تحقیقاتی سیستمهای دانش محاسبه شد [ 32 ، 33 ].
نقشه های طبقه بندی سه مرحله زمانی گسترش جنگل به صورت پیکسل به پیکسل با پارامترهای خاک و توپوگرافی مقایسه شد و برای هر مورد Kloc محاسبه شد.
2.5. شرح ساختار جنگل فعلی
علاوه بر تغییرات مکانی و زمانی بر اساس ماتریسهای انتقال، ترکیب گونههای درختی و ساختار جنگل را در تودههای مختلف با توجه به سن جنگل برای سال 2008 شناسایی کردیم. در هر یک از سایتهای مطالعه ما، شبکهای از 25 نقطه با فاصله منظم در هر 200 متر به طور تصادفی روی نقشه قرار گرفته است. در هر نقطه، پوشش گیاهی را بر اساس روشی که براون-بلانکه [ 34 ] توسعه داده بود بررسی کردیم و ارتفاع افراد چوبی را روی سطحی که با توجه به تراکم درخت متفاوت بود، اندازه گیری کردیم، یعنی 64 متر مربع برای جنگل ها، جایی که پوشش جنگلی است. درختان کمتر از 10٪ است و 200 متر مربع برای مناطق جنگلی، که در آن پوشش درختان جنگلی بیشتر از 10٪ است [ 35 ]].
3. نتایج
3.1. ارزیابی دقت طبقه بندی
دقت کلی نقشه پوشش گیاهی در سال 2003 0.84 است، با 63 پیکسل به همین ترتیب توسط تفسیر عکس و بررسی زمین طبقه بندی شده است. خطاها عمدتاً شامل جنگل های مختلط است که در اینجا بیش از حد برآورد شده است. اینها را می توان با تفاوت سطح تحلیل شده بین عکس های میدانی و هوایی توضیح داد.
3.2. دینامیک منظره
مقایسه سطوح با دستههای مختلف پوشش گیاهی در طول سالها از سال 1946 نشان میدهد که در طول زمان با افزایش نرخ پیشرفت در طول این دورهها کاهش در مناطق گرمسیری را نشان میدهد ( شکل 3 a,b را ببینید). بیش از 60 سال، بیش از 75 درصد از گرمسیری به تدریج توسط گونه های چوبی مستعمره شده است ( جدول 2 را ببینید ). این تغییر ناهمگونی مکانی-زمانی مشخصی را در هر یک از سه مکان میدانی ما ایجاد کرده است همانطور که در شکل 4 نشان داده شده است.
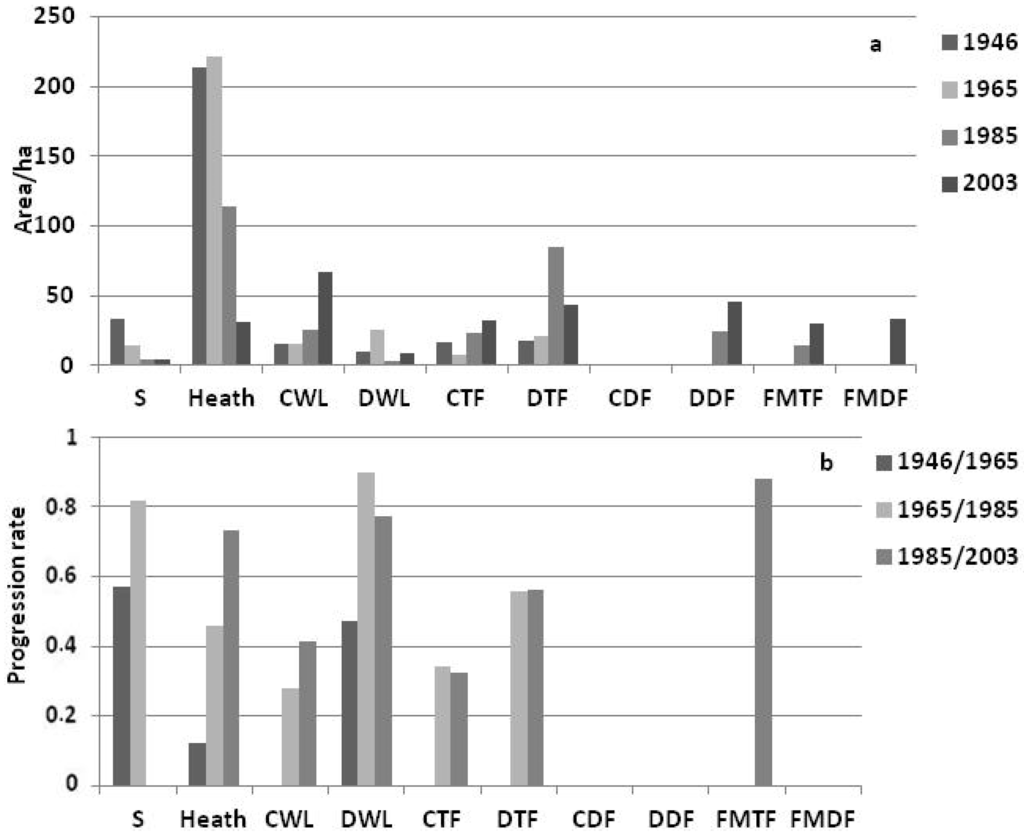
شکل 3. ( الف ) تغییرات در مساحت (هکتار) مراحل مختلف دینامیکی بین سالهای 1946 و 2003. ( ب ) تغییرات در نرخ تغییر برای هر دسته پوشش گیاهی محاسبه شده از ماتریسهای گذار 1946-1965، 1965-1985، و 1985-1985 2003 (به عنوان مثال، نرخ تغییر هیتلند 0.13 بین سال های 1946 و 1965 و 0.73 بین سال های 1985 و 2003 است). خاک لخت (S)، Heathland (Heath)، جنگل مخروطیان (CWL)، جنگل های برگریز (DWL)، جنگل نازک مخروطیان (CTF)، جنگل نازک برگریز (DTF)، جنگل انبوه مخروطیان (CDF)، جنگل انبوه برگریز (DDF) , جنگل نازک جنگلی مختلط (FMTF), جنگل انبوه جنگلی مختلط (FMDF).
جدول 2. ماتریس های انتقال با درصد تغییر از یک مرحله به مرحله دیگر بین دو تاریخ: ( الف ) 1946-1965، ( ب ) 1965-1985، و ( ج ) 1985-2003. پسزمینه خاکستری تیره نشاندهنده گذار پیشرونده هولند به جنگل است، پسزمینه خاکستری روشن نشاندهنده گذار قهقرایی از جنگل به گرمسیری مربوط به فعالیتهای انسانی است، و مقادیر زیر خطدار نشاندهنده خط مورب بدون تغییر پوشش گیاهی مختلف است. دسته بندی پوشش گیاهی در جدول 1 توضیح داده شده است.
خاک برهنه عمدتاً توسط گونههای گرمسیری، یعنی 32 درصد در بازه زمانی اول و 52 درصد در بازه دوم استعمار شدند، در حالی که استعمار خاک لخت توسط گونههای چوبی کمتر بود، یعنی 16 درصد و 26 درصد در نوبت اول و دوم. فواصل، به ترتیب؛ این عمدتاً به دلیل استعمار جوامع جنگلی برگریز بود ( جدول 2 a,b را ببینید). در طول دو دوره اول مورد مطالعه، 1946-1965 و 1965-1985، مناطق گرمسیری توسط جوامع برگریز DWL، DTF و DDF به ترتیب بیش از 9٪ و 29٪ از سطح مستعمره شدند، در حالی که استعمار هولند توسط مخروطیان CWL، CTF کندتر بود. به ترتیب 4% و 15% را تشکیل می دهند ( جدول 2 را ببینیدالف، ب). با این حال، در طول دوره نهایی، 1985-2003، استعمار جنگل سریعتر بود و اساساً به دلیل درختان مخروطی CWL، CTF، یعنی 53٪ در مقابل جوامع برگریز DWL با 7٪ و گونه های جنگلی مخلوط با 13٪ ( جدول 2 c). Heathland مستعمره جوامع برگریز (DWF) یک مرحله سریع از تکامل را بین سالهای 1965 و 1985 نشان داد. 76 درصد از مساحت از DWF به DTF تغییر کرد ( جدول 2 ب).
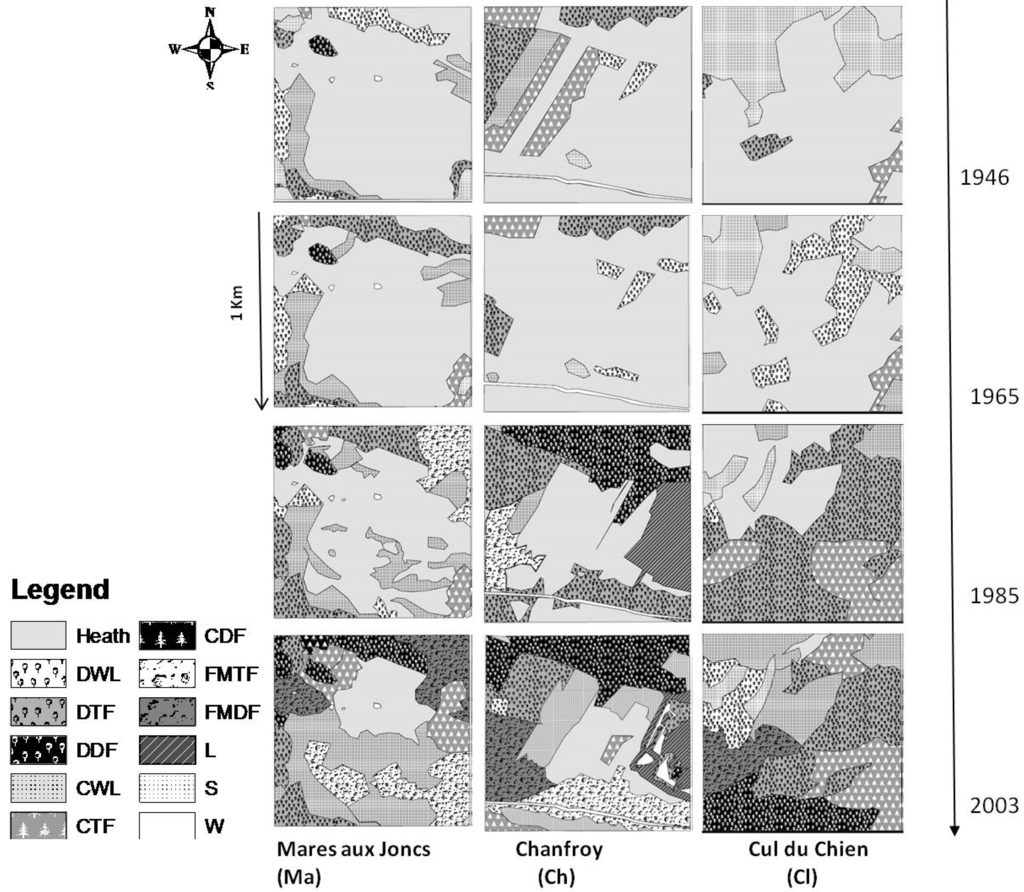
شکل 4. نقشههای زیستگاه برای سالهای 1946، 1965، 1985، و 2003. پالت مقیاس خاکستری مورد استفاده برای ارائه نقشهها، منعکسکننده پیشرفت توالی ثانویه بین مناطق باز (رنگهای روشن) و جنگلها (رنگهای تیرهتر) است. خاک لخت (S)، Heathland (Heath)، جنگل مخروطیان (CWL)، جنگل های برگریز (DWL)، جنگل نازک مخروطیان (CTF)، جنگل نازک برگریز (DTF)، جنگل انبوه مخروطیان (CDF)، جنگل انبوه برگریز (DDF) , جنگل نازک جنگلی مختلط (FMTF)، جنگل انبوه جنگلی مختلط (FMDF)، چمن (L)، آب (W).
جنگل های احیا شده توسط درختان مخروطی در طول دوره 1946-1965 پایدار باقی ماندند. بیست و هشت درصد از سطح به سمت جنگل نازک مخروطیان و جنگل های مختلط نازک در طول دوره 1965-1985 تغییر کرد. این نسبت طی دوره 1985-2003 به 42% افزایش یافت ( شکل 3 ب و جدول 2 ). جنگلهای مخروطی نازک عمدتاً به جنگلهای مختلط یا برگریز تغییر یافتهاند، به ویژه در مناطقی که مستقیماً در مجاورت مناطق جنگلی خزانریز قرار دارند ( شکل 4 ). تغییر مهمی در پوشش جنگلی در این دوره نیز مشاهده شد ( شکل 5 د). همانطور که در شکل 5 نشان داده شده است، افزایش جنگل های انبوه برگریز و جنگل های انبوه مخلوط را به قیمت جنگل های نازک مشاهده کردیم .قبل از میلاد مسیح. با این حال، مساحت جنگل های مخروطی نازک افزایش یافت، در حالی که تغییر در جنگل های انبوه مخروطیان تقریباً صفر بود ( شکل 5 الف را ببینید). بر این اساس، ما می توانیم به وضوح بسته شدن فضاهای باز را با شتاب این پویایی در طول زمان مشاهده کنیم.

شکل 5. ( الف ) تغییرات در مساحت (هکتار) درختان مخروطی بین سال های 1946 و 2003: جنگل های مخروطی (CWL)، جنگل های نازک مخروطیان (CTF)، جنگل های انبوه مخروطیان (CDF). ( ب ) تغییرات در مساحت (هکتار) برگریزان بین سالهای 1946 و 2003: جنگلهای برگریز (DWL)، جنگلهای نازک برگریز (DTF)، جنگلهای انبوه برگریز (DDF). ( ج ) تغییرات در مساحت (هکتار) جنگل مختلط بین سالهای 1946 و 2003: جنگل نازک جنگلی مختلط (FMTF)، جنگل انبوه جنگلی مختلط (FMDF). ( د ) تغییرات در مساحت (هکتار) انواع مختلف جنگل: برگریز (D)، مخروطیان (C)، جنگل مختلط (FM) بین سالهای 1946 و 2003.
3.3. تأثیر متغیرهای محیطی فضایی بر پویایی جنگل
نتایج نشان داد که پویایی جنگل مستقل از نوع خاک ( P <0.0001)، عمق خاک ( P <0.05)، یا جهت مواجهه آنها ( P <0.05) نبود. با این حال، هیچ تاثیری از شیب بر پویایی جنگل وجود نداشت. Heathland بدون توده های استعمار درخت از نظر فضایی با خاک کم عمق (Kloc = 0.56)، خاک اسیدی و ماسه سنگ (Kloc = 0.5)، و توپوگرافی مسطح (Kloc = 0.6) مرتبط بود. به تازگی استعمار هیتلند می ایستد، به عنوان مثال، 1985-2003، همچنین مربوط به خاک اسیدی و ماسه سنگ (Kloc = 0.4)، خاک های با عمق متوسط (Kloc = 0.4)، و جنبه جنوب (Kloc = 0.39) بود. استعمارهای اولیه اولیه (1965-1985) درجه قابل توجهی از توافق با خاک عمیق (Kloc = 0.52)، luvisols اسیدی (Kloc = 0.43)، و یک جنبه رو به شمال (Kloc = 0.6) را نشان داد. استعمار گونههای درختی بین سالهای 1946 و 1965 ترجیحاً روی لوویزولهای با عمق متوسط و اسیدی اتفاق افتاد (Kloc = 0.42). هیچ ترجیح فضایی دیگری برای این پایه تشخیص داده نشد.
3.4. ساختار جنگل و ترکیب گونه ها
اشغال کاج ( Pinus sylvestris L.) در تودههای خالص در مناطقی که اخیراً جنگلکاری شدهاند، به عنوان مثال ، در دوره 20 ساله اخیر بیشتر است، در حالی که تودههای مخلوط Pinus sylvestris و Betula pendula Roth. در مناطقی که بین 20 تا 40 سال پیش جنگل کاری شده اند غالب هستند. تودههای کاج بلوط متشکل از Pinus sylvestris و Quercus robur L. در مناطقی که در طول 40 سال گذشته جنگلکاری شدهاند غالب هستند، در حالی که آن مناطقی که جنگلهای قدیمیتری دارند عمدتاً توسط دو جامعه اشغال شدهاند: Q. robur ، Castanea sativa Mill.، P. سیلوستریس ، Pinus pinaster Aiton.، وQ. robur ، B. pendula ، و P. sylvestris . اندازه گیری ارتفاع درختان منفرد از 11 متر تا 20 متر برای مناطق جنگل کاری شده از 1946 تا 1965 (60 سال پیش)، 9 تا 17 متر در مناطق جنگل کاری شده از 1965 تا 1985 (40 سال پیش) و 7 متر تا 14 متر متغیر بود. در مناطق جنگلی شده از 1985-2003 (تا 20 سال پیش).
4. بحث
تغییرات چشم انداز مشاهده شده در این دوره با تغییر شدید استفاده از زمین توسط انسان مطابقت دارد. تا اوایل دهه 1960 این منطقه از جنگل مالکیت خصوصی داشت که ناپدید شدن جنگل های چوبی بین سال های 1946 و 1965 به دلیل فعالیت های انسانی را توضیح می دهد. از جمله فعالیتهای تاریخی در این منطقه، معادن قدیمی در چانفروی نیز میباشند که نمایانگر ظهور علفزارها در این مکان در دوره 1965-1985 است که نتیجه بازسازی معادن است.
جانشینی پوشش گیاهی طبیعی پس از توقف فعالیتهای سنتی کشاورزی – مرتعی و بهرهبرداری از جنگلها دوباره آغاز شد. در دو دوره اولی که مورد مطالعه قرار گرفتیم، یعنی بین سالهای 1946 و 1985، ما گسترش گرمابه را به ضرر خاک لخت تشخیص دادیم ( جدول 2 a,b)، در حالی که استعمار خاکهای لخت توسط گونههای چوبی در این دوره کمتر بود. استعمار توسط Calluna vulgaris [ 36 ]، یک گونه جانشین اولیه [ 34 ]، توسط شرایط ادافیک محلی مورد علاقه بود: خاکهای شنی و اسیدی که استعمار توسط سایر گونه های چوبی را دشوار می کند.
ما شتابی را در استعمار گونههای چوبی در طول زمان مشاهده کردیم. Heathland که برای طولانی ترین دوره از نفوذ انسان حذف شده بود، بیشترین احتمال را داشت که توسط گونه های چوبی مستعمره شود. دو توضیح برای این افزایش در طول زمان می توان ارائه کرد. اول، پیری هیتلند مربوط به پایان چرخه زندگی C. vulgaris است که حدود 35 سال تخمین زده می شود [ 37 ]. در این زمان هالند وارد مرحله انحطاط شد که مربوط به هجوم گونه های چوبی در خاکی مهمان نوازتر بود. رقابت با گونه های چوبی مانع از شروع چرخه زندگی Calluna شد [ 38] و بنابراین استعمار گونه های چوبی در فضای آزاد موجود را مورد علاقه قرار داد. ثانیاً، در ابتدای گسترش جنگل، یک جبهه استعمار در اطراف منطقه جنگلی موجود آغاز شد ( شکل 5 ) و این با بسته شدن مناطق جنگلی موجود همراه بود ( جدول 2 ). با این حال، استعمار مستقیم گونههای چوبی در مناطق باز در طول دوره اخیر در پویایی بیشتر بود ( شکل 4 را ببینید )، و این باعث افزایش سرعت جنگلکاری هاتلها شد. چنین پویایی فضایی با مفهوم کلاسیک استعمار مجدد توسط گونه های چوبی مطابقت دارد [ 39 ، 40 ]]، نشان میدهد که گونههای چوبی پیشگام انیموکرو و هلیوفیل دو استراتژی استعمار دارند: یا با پراکندگی با استعمار مستقیم در محیطهای باز، یا با گسترش در مجاورت یک جنگل مستقر. در سایت های مطالعه ما، P. sylvestris یکی از دو گونه چوبی اصلی پیشگام بود. Debussche & Lepart [ 41 ] و Debain [ 42]، کار بر روی این گونه نشان داد که جبهه اصلی استعمار آن در چند ده متری اطراف تودههای کاج قدیمی قرار داشت و افرادی که پیش از جبهه استعمار تودههای کاج قدیمی مستقر شدهاند، فضای اطراف آنها را مستعمره میکنند. در اینجا، بلوغ جمعیت های چوبی که قبل از سال 1946 وجود داشتند، امکان استعمار هولند را نسبتاً سریع فراهم کرد. استعماری که از جلو توسط افراد پراکنده یا افراد منزوی گسترش مییابد با زمان لازم برای رسیدن افراد به سن باروری کند میشود. این بدان معناست که 15 سال یا بیشتر برای P. sylvestris [ 43 ] و 20 سال برای B. pendula [ 44 ] است.] قبل از اینکه این افراد پراکنده بتوانند استعمار اطراف خود را آغاز کنند که منجر به بسته شدن تدریجی پوشش گیاهی می شود.
درک استعمارهای دیفرانسیل هولند رها شده توسط گونه های برگریز و مخروطیان اجازه می دهد تا فرآیند پیچیده جانشینی طبیعی و پویایی آن مورد تجزیه و تحلیل قرار گیرد. پویایی گونه های برگریز در طول دوره 1965-1985 تا حدی از بسته شدن مناطق جنگلی باز توسط گونه های پیشگام برگریز و تا حدودی از استعمار فضاهای باز ناشی می شود ( شکل 4 ب و جدول 2 ب را ببینید). برای دوره 1985-2003، پویایی گونه های برگریز اساساً به بسته شدن مناطق چوبی موجود محدود شد. ما متوجه شدیم که استعمار گونه های چوبی در این دوره اساساً توسط گونه های مخروطی، با افزایش قابل توجهی در سطح اشغال شده توسط کاج است. این گسترش سوزنی برگ در سایت دیگری در این منطقه نیز مشاهده شده است [11 ].
چندین توضیح را می توان برای توضیح استعمار توسط درختان مخروطی در طول دوره 1985-2003 ارائه کرد. اول، تداوم فضایی این استعمار را می توان با ظرفیت مخروطیان برای استعمار و رشد در خاک های کم عمق و فقیر که در آن گونه های پیشگام برگریز قادر به ایجاد جای پایی نیستند توضیح داد ( شکل 5 a). P. sylvestris قادر است خاکهای فقیر و خشک را مستعمره کند [ 43 و 45]. جنگلکاری در این دوره میتواند به استعمار گونههای مخروطی در مناطقی که برای گونههای برگریز نامطلوب است مرتبط باشد، مساعدترین مناطق قبلاً توسط گونههای برگریز مستعمره شدهاند. این فرضیه از طریق نتایج خاک سنجی تأیید شده است. ما در این دوره با توجه به شرایط خاک متوجه دو مرحله استعمار گونههای چوبی شدیم: لوویزولهای حاصلخیز و خاک عمیق. فاز اول با استعمار مناطق مساعد توسط گونه های برگریز و بسته شدن پوشش گیاهی جنگل مشخص می شود و به تدریج یک جنگل بالغ را تشکیل می دهد. در مرحله دوم، گونه های مخروطی در مناطقی که شرایط خاک برای نصب گونه های برگریز نامناسب است، مستعمره می شوند .، خاک های فقیر و کم عمق مانند خاک اسیدی و ماسه سنگ.
ثانیا، گرم شدن سریع آب و هوا که از دهه 1980 به بعد در منطقه کلان شهر فرانسه ثبت شد [ 46 ، 47 ] و تغییرات بارش فصلی، به ویژه کاهش بارندگی تابستان [ 47 ، 48 ]، منجر به دوره ای از خشکسالی شده است که ممکن است منجر به خشکسالی شود. یک مزیت انتخابی برای P. sylvestris به دلیل نیاز آبی کم و تنظیم مطلوب روزنهای [ 49 ] بود. ثالثاً، این تغییرات آب و هوایی ممکن است پیامدهایی برای فنولوژی و مدت زمان فصل پوشش گیاهی داشته باشد. منزل و فابیان [ 50] از دهه 1960 طولاني شدن فصل رشد فعال را نشان داده اند. این پدیده می تواند با افزایش تولید سالانه فتوسنتزی درختان سوزنی برگ به دلیل توانایی آنها در استفاده از تابش خورشیدی موجود در زمستان و اوایل بهار برای آنها مفید باشد.
در میان عوامل مورد مطالعه در اینجا، انواع خاک و عمق خاک تا حد زیادی جهت پویایی جنگل را تعیین می کند. با توجه به تأثیر متغیرهای فیزیوگرافیک بر پویایی جنگل، جهت گیری دامنه ها بر پویایی پوشش گیاهی تأثیر دارد، به طوری که ما تداوم گرمسیری را در دامنه های گرمتر جنوب رو به جنوب که مناسب تر برای پوشش گیاهی گرمک خشک هستند مشاهده کردیم. دامنههای شمالی مرطوبتر هستند و استعمار درختان و سایر درختچهها در آنجا آشکارتر است. هیچ تأثیری از شیب زمین (شیب شیب) تشخیص داده نشد، که می توان با این واقعیت توضیح داد که شیب های تند زمینی حاشیه ای برای کشاورزی هستند و قبل از سال 1946 رها شده بودند، سالی که پایش دینامیکی در مطالعه ما آغاز شد.
استفاده از عکسهای هوایی برای ایجاد نقشههای اکوسیستم، فرآیندی بدون خطا نیست. خطاهای کمی و/یا کیفی ممکن است، ناشی از دیجیتالی کردن، ارجاع جغرافیایی، اندازه گیری، طبقه بندی و خطاهای درون یابی [ 51 ]. به همین دلیل عدم قطعیت های مکانی در داده های تاریخی ورودی ما به دلیل عدم وجود داده های زمینی قابل برآورد نیست و انتشار خطا مربوط به عدم قطعیت ورودی ارزیابی نشده است.
5. نتیجه گیری و پیامدها برای آمایش سرزمین
مطالعه ما استعمار مجدد مناطق گرمسیری توسط جنگل ها را با از دست دادن بیش از 75 درصد از مناطق گرمسیری طی 60 سال گذشته مستند می کند. با این وجود، این استعمار از نظر زمانی و مکانی متفاوت است، زیرا فرآیند استعمار مجدد جنگل به عوامل متعددی بستگی دارد: مکانیسمهای زیربنایی پویایی گونههای چوبی، سولههای اکولوژیکی گونههای مختلف، و تنوع فضایی در خاک و شرایط اقلیمی در داخل سایت.
ارتباط این مطالعه برجسته کردن اهمیت در نظر گرفتن عوامل مکانی و زمانی برای مدیریت حفاظت از گرمسیری است. این مفهوم در هیچ مطالعه قبلی در مورد مدیریت گرمسیری اعمال یا پیشنهاد نشده است.
نتایج ما نشان میدهد که عامل زمانی بسیار مهم است، زیرا هلند با سرعت یکسان در طول زمان مستعمره نمیشود. در نظر گرفتن این اطلاعات، همراه با عوامل فضایی ارجح برای تداوم گرمسیری، می تواند یک مزیت بزرگ برای مدیریت گرمسیری باشد. همچنین این پتانسیل را دارد که با تمرکز بر مناطق و شرایط فضایی که از احیای جنگل ها جلوگیری می کند و از این رو به پایداری درازمدت گرمسیری کمک می کند، به مدیران زمین در انتخاب مناطقی که حفاظت و/یا احیای آن ها آسان تر است، کمک کند. با توجه به نتایج ما، اولویت مدیریت باید به آن دسته از گرمکهایی داده شود که در خاکهای کمعمق، در سطوح هموار، یا سایتهای رو به جنوب و دور از مرزهای جنگلهای بالغ قرار دارند.
منابع
- فولی، جی. دفریز، ر. آسنر، GP; بارفورد، سی. بونان، جی. نجار، SR; چاپین، FS; Coe, MT; روزانه، GC; گیبس، هنگ کنگ؛ و همکاران پیامدهای جهانی استفاده از زمین Science 2005 ، 309 ، 570-574. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- راکستروم، جی. استفن، دبلیو. Noone، K. پرسون، Å. Chapin، FS، III; لامبین، EF; لنتون، TM; شفر، ام. فولک، سی. Schellnhuber، HJ; و همکاران فضای عملیاتی امن برای بشریت. طبیعت 2009 ، 24 ، 472-475. [ Google Scholar ]
- دستورالعمل EC Habitats. دستورالعمل 92/43/EEC شورای 21 مه 1992 در مورد حفاظت از زیستگاه های طبیعی و جانوران و گیاهان وحشی . کمیسیون اروپا: بروکسل، بلژیک، 1992. [ Google Scholar ]
- Ausden، M. مدیریت زیستگاه برای حفاظت. کتابچه راهنمای تکنیک ها ; انتشارات دانشگاه آکسفورد: آکسفورد، انگلستان، 2007; پ. 384. [ Google Scholar ]
- تامپسون، DBA; مک دونالد، ای جی؛ Marsden، JH; گالبریت، کالیفرنیا مرتع هدر بالادست در بریتانیای کبیر: مروری بر اهمیت بینالمللی، تغییر پوشش گیاهی و برخی اهداف برای حفاظت از طبیعت. Biol. حفظ کنید. 1995 ، 71 ، 163-178. [ Google Scholar ] [ CrossRef ]
- Usher, MB مدیریت و تنوع بندپایان در Calluna Hethland. تنوع زیستی حفظ کنید. 1992 ، 1 ، 63-79. [ Google Scholar ] [ CrossRef ]
- آشر، مگابایت؛ تامپسون، تنوع DBA در کوهستان های مرتفع بریتانیای کبیر: اهمیت حفاظت. Biol. حفظ کنید. 1993 ، 66 ، 69-81. [ Google Scholar ] [ CrossRef ]
- وب، NR مدیریت سنتی مناطق گرمسیری اروپا. J. Appl. Ecol. 1998 ، 35 ، 987-990. [ Google Scholar ] [ CrossRef ]
- وب، هلند شمال اقیانوس اطلس. در کتاب دیوی احیای محیط زیست ; Martin, RP, Anthony, J., Eds. انتشارات دانشگاه کمبریج: کمبریج، انگلستان، 2008; جلد 1 و 2، ص 401-418. [ Google Scholar ]
- قیمت، EAC دشت مرتع و زیستگاه Heathland ; Routledge: لندن، انگلستان، 2003. [ Google Scholar ]
- مبعید، س. ریرا، بی. لالان، ا. باگت، م. Machon، N. استفاده از رویکردهای فضایی دیاکرونیک و مدلسازی پیشبینیکننده برای مطالعه پویایی پوشش گیاهی یک هیولند مدیریت شده. تنوع زیستی حفظ کنید. 2011 ، 20 ، 73-88. [ Google Scholar ] [ CrossRef ]
- بارکر، سی جی; پاور، SA; بل، JNB؛ Orme، CDL اثرات مدیریت زیستگاه بر پاسخ دهلیز به رسوب نیتروژن اتمسفر. Biol. حفظ کنید. 2004 ، 120 ، 41-52. [ Google Scholar ] [ CrossRef ]
- Wamelink، GWW؛ دی جونگ، جی جی. ون دوبن، HF; van Wijk، MN هزینه های اضافی مدیریت طبیعت ناشی از رسوب. Ecol. اقتصاد 2005 ، 52 ، 437-451. [ Google Scholar ] [ CrossRef ]
- کالوو، ال. آلونسو، آی. مارکوس، ای. د لوئیس، E. اثرات برش و رسوب نیتروژن بر تنوع زیستی در کانتابریا. Appl. گیاهی علمی 2007 ، 10 ، 43-52. [ Google Scholar ] [ CrossRef ]
- Fagundez، J. Heathlands در مواجهه با تغییرات جهانی: محرک های از دست دادن تنوع زیستی از سناریوهای گذشته تا آینده. ان ربات 2013 ، 111 ، 151-172. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Miles, J. Vegetation Dynamics ; چپمن و هال: لندن، بریتانیا، 1979. [ Google Scholar ]
- ایوانز، اف سی؛ دهل، ای. ساختار رویشی یک مزرعه متروک در جنوب شرقی میشیگان و روابط آن با عوامل محیطی. اکولوژی 1955 ، 36 ، 685-706. [ Google Scholar ] [ CrossRef ]
- Dargie، TCD تجزیه و تحلیل ترتیب الگوهای پوشش گیاهی بر روی شیب های آب و هوا در جنوب شرقی اسپانیا. J. Biogeogr. 1987 ، 14 ، 197-211. [ Google Scholar ] [ CrossRef ]
- بادانو، EI; کاویرس، لس آنجلس؛ مولینا-مونته نگرو، MA; Quiroz، جنبه شیب CL بر الگوهای ارتباط گیاهی در منطقه مدیترانه ای مرکزی شیلی تأثیر می گذارد. J. محیط خشک. 2005 ، 62 ، 93-108. [ Google Scholar ] [ CrossRef ]
- تیلمن، دی. Kareiva، P. اکولوژی فضایی: نقش فضا در پویایی جمعیت و تعاملات بین گونه ای . دانشگاه پرینستون: پرینستون، نیوجرسی، ایالات متحده آمریکا، 1997; پ. 368. [ Google Scholar ]
- مبعید، س. پونگ، جی اف. سالمون، اس. لالان، ا. Riera، B. تأثیر تغییرپذیری فضایی نوع خاک و استعمار درخت بر پویایی Molinia caerulea ( L. ) Moench در هتک مدیریت شده. Ecol. مجتمع. 2012 ، 11 ، 118-125. [ Google Scholar ] [ CrossRef ]
- جیمز، SA; ایرنه، م. Johannes, R. Small Format Aerial Photography: Principles, Techniques and Geoscience Applications ; Elsevier Science: Trier، آلمان، 2010; پ. 268. [ Google Scholar ]
- دریر، ن. Lucas, S. La Forêt Française en 2005 Results de la Première Campagne Nationale Annuelle ; IFN: Saint-Mandé، فرانسه، 2006; پ. 8. [ Google Scholar ]
- راس، اس ال. جان، سنجش از دور GL و ارزیابی دقت GIS ; آژانس حفاظت از محیط زیست ایالات متحده (EPA): واشنگتن، دی سی، ایالات متحده آمریکا، 2011; پ. 320.
- ایدریسی ، نسخه آند 32; دانشگاه کلارک: Worcester، OH، ایالات متحده آمریکا، 1987-2006.
- ESRI ArcGIS نسخه 9.2. Environmental Systems Research Institute Inc.: Redlands، CA، USA، 2006.
- ایزاکس، EH; Srivastava, RM مقدمه ای بر زمین آمار کاربردی ; انتشارات دانشگاه آکسفورد: نیویورک، نیویورک، ایالات متحده آمریکا، 1989; پ. 561. [ Google Scholar ]
- کریگ، دی. یک مسئله ایستا برای برخی از مشکلات اساسی ارزش گذاری حداقل در Witwatersrand. جی. شیمی. فلزی. حداقل Soc. اس افر. 1951 ، 52 ، 119-139. [ Google Scholar ]
- موسسه جغرافیایی ملی (فرانسه). نقشه خاک Forêt de Trois Pignons ONF/ED EDR25©IGN2006 [نقشه] ; گزارش فنی Forêt Domaniale des Trois Pignons; Office National des Forêts: پاریس، فرانسه، 2006.
- خطای Pontius، RG Quantification در مقابل خطای مکان در مقایسه نقشه های طبقه بندی شده. فتوگرام مهندس Remote Sens. 2000 , 66 , 1011–1016. [ Google Scholar ]
- لندیس، جی آر. Koch, GG اندازه گیری توافق ناظر برای داده های طبقه بندی شده. بیومتریک 1977 ، 33 ، 159-174. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- ویسر، اچ. de Nijs, T. کیت مقایسه نقشه نرم افزار MCK. محیط زیست مدل نرم افزار 2006 ، 21 ، 346-358. [ Google Scholar ] [ CrossRef ]
- کیت مقایسه نقشه 3. موجود به صورت آنلاین: www.riks.nl (در 21 فوریه 2014 قابل دسترسی است).
- براون بلانکه، جی. جامعه شناسی گیاهی . McGraw-Hill Book Company: نیویورک، نیویورک، ایالات متحده آمریکا، 1932; پ. 438. [ Google Scholar ]
- میلان، سی. Zdenka، O. اندازههای قطعه مورد استفاده برای نمونهبرداری جامعهشناسی گیاهی از پوشش گیاهی اروپا. J. Veg. علمی 2003 ، 14 ، 563-570. [ Google Scholar ]
- گیمینگهام، CH اکولوژی هیتلند ؛ چپمن و هال: لندن، انگلستان، 1972; پ. 266. [ Google Scholar ]
- واکر، ال آر. واکر، جی. هابز، RJ پیوند مرمت و جانشینی زیست محیطی . Springer: نیویورک، نیویورک، ایالات متحده آمریکا، 2007. [ Google Scholar ]
- Gimingham، CH ارزیابی مجدد فرآیندهای چرخه ای در Calluna Hethland. گیاهی 1988 ، 77 ، 61-64. [ Google Scholar ] [ CrossRef ]
- گریم، جی پی؛ هاجسون، جی جی. هانت، آر. بومشناسی مقایسهای گیاهی: رویکردی کاربردی به گونههای رایج بریتانیایی . Unwin Hyman: لندن، انگلستان، 1988; پ. 742. [ Google Scholar ]
- رامو، جی سی. عمارت، دی. Dumé, G. Flore Forestière Française ; Institut pour le Développement Forestier: Dijon، فرانسه، 1993; پ. 2421. [ Google Scholar ]
- دبوشه، ام. Lepart, J. استقرار گیاهان چوبی در مزارع قدیمی مدیترانه: فرصت در فضا و زمان. Landsc. Ecol. 1992 ، 6 ، 133-145. [ Google Scholar ] [ CrossRef ]
- دیبین، اس. کرت، تی. لپارت، جی. Prevosto، B. تنوع تولید مثل در Pinus sylvestris در جنوب فرانسه: پیامدهایی برای تهاجم. J. Veg. علمی 2003 ، 4 ، 509-516. [ Google Scholar ] [ CrossRef ]
- ریچاردسون، DM اکولوژی و جغرافیای زیستی کاج ; انتشارات دانشگاه کمبریج: کمبریج، انگلستان، 1988; پ. 546. [ Google Scholar ]
- شرر-لورنزن، ام. کرنر، سی. Schulze، ED سیستم های معتدل و شمالی. در تنوع و عملکرد جنگل ; Springer: برلین، آلمان، 2005; صص 377-390. [ Google Scholar ]
- کانیلاس، آی. مارتینز گارسیا، اف. Montero، G. جنگلکاری و پویایی کاج سیلوستریس L. در اسپانیا. تحقیق کنید. آگرار. سیست عود کند. برای. 2000 ، 1 ، 233-253. [ Google Scholar ]
- بسمولین، پی. Mestre, O. Le réchauffement climatique sur le siècle en فرانسه. Fr. چانگ. گلوب. 2001 ، 12 ، 32-34. [ Google Scholar ]
- کانلاس، سی. Gibelin، AL; Lassègues، P. کردونکاف، م. دندن، پ. Simon, P. Les normales climatiques spatialisées Aurelhy 1981-2010: دما و بارش. La Météorol. 2014 ، 85 . [ Google Scholar ] [ CrossRef ]
- Moisselin، JM Les précipitations en France au XXème siècle. Fr. چانگ. گلوب. 2002 ، 13 ، 57-62. [ Google Scholar ]
- دولامسورن، سی. هاک، ام. بادر، م. اویونگرل، اس. اسوخجارگل، د. نیامبایار، س. Leuschner, C. استراتژیهای مختلف Pinus sylvestris و Larix sibirica برای مقابله با خشکسالی تابستانی در شمال مغولستان جنگلی-استپی اکوتون، برتری آینده کاج را در آب و هوای گرمتر نشان میدهد. می توان. جی. برای. Res. 2009 ، 39 ، 2520-2528. [ Google Scholar ] [ CrossRef ]
- منزل، ع. Fabian, P. فصل رشد در اروپا تمدید شد. طبیعت 1999 ، 397 ، 659-661. [ Google Scholar ] [ CrossRef ]
- روچینی، دی. Foody, DG; ناژندرا، اچ. ریکوتا، سی. آناند، م. او، KS; آمیسی، وی. کلاینشمیت، بی. فورستر، ام. اشمیتلین، اس. و همکاران عدم قطعیت در نقشه برداری اکوسیستم با سنجش از دور محاسبه کنید. Geosci. 2013 ، 50 ، 128-135. [ Google Scholar ] [ CrossRef ]
© 2015 توسط نویسندگان; دارنده مجوز MDPI، بازل، سوئیس. این مقاله یک مقاله با دسترسی آزاد است که تحت شرایط و ضوابط مجوز Creative Commons Attribution (http://creativecommons.org/licenses/by/4.0/) توزیع شده است.
بدون نظر