1. معرفی
روشهای آماری مورد استفاده برای ارتباط تنوع زیستی با عوامل محیطی برای پرداختن به مجموعهای از پرسشهای اکولوژیکی از پیشبینی اثرات تغییرات آب و هوایی بر تنوع زیستی [ 1 ]، شناسایی تغییرات انسانی پوشش زمین بر غنای گونهها [ 2 ] تا کاوش مکانهای جدید ضروری هستند. سایت های رزرو [ 3 ]. مطالعات سنتی که تنوع زیستی را بررسی میکنند اغلب رابطه بین غنای گونهای (تعداد گونههای مختلف ثبتشده در یک منطقه) و مجموعهای از عوامل محیطی را در یک مقیاس فضایی واحد بررسی کردهاند [ 4 ، 5 ، 6 ، 7 ]]. انتخاب مقیاس فضایی هنگام مطالعه تنوع زیستی بهویژه مهم است، زیرا عوامل و فرآیندهایی که در یک مقیاس مهم هستند اغلب در مقیاس دیگر اهمیت کمتری دارند و تفسیر و پیشبینی را دشوار میکنند [ 8 ]. بسیاری از مطالعات متعاقباً یک رویکرد چند مقیاسی را برای مطالعه فرآیندهای مسئول الگوهای فضایی مشاهده شده در تعدادی از مقیاسهای فضایی اتخاذ کردهاند [ 9 ، 10 ، 11 ].
اخیراً، آمار فضایی محلی برای بررسی الگوهای غنای گونهای در رابطه با خودهمبستگی فضایی [ 12 ، 13 ، 14 ] و غیرایستایی فضایی [ 15 ، 16 ، 17 ، 18 ، 19 استفاده شده است.]. یکی از مهمترین پارامترها در آمار فضایی محلی، ماتریس وزن های فضایی است که برای تعیین مشاهدات “همسایه” استفاده می شود. محله ها عموماً بر اساس فاصله (آستانه ها یا وزن دهی معکوس فاصله) یا مجاورت هستند و انتخاب تعریف محله مناسب اغلب ذهنی است با اکتشاف داده های کمی که از قبل انجام شده است. بنابراین، مطالعاتی که به بررسی تأثیر عوامل محیطی بر غنای گونهای میپردازند، باید چگونگی تأثیر مقیاس فضایی بر روابط غنا-محیط مشاهده شده و همچنین روشهای آماری مورد استفاده برای مطالعه آنها را در نظر بگیرند.
خودهمبستگی فضایی به رابطه بین شباهت و فاصله اشاره دارد، که چیزهای نزدیک بیشتر از چیزهای دور مرتبط هستند، و مبنای قانون اول جغرافیای توبلر [ 20 ] را فراهم می کند. این پدیده اغلب در دادههای گونهها مشاهده میشود، و میتواند ناشی از فشارهای زیستی، مانند رقابت یا پراکندگی، و همچنین از ترجیحات زیستگاه در شیبهای محیطی ساختار فضایی [ 21 ] باشد. اخیراً تأثیر آماری خودهمبستگی فضایی در مطالعات بومشناختی مورد توجه قرار گرفته است (نگاه کنید به [ 22 ، 23 ، 24 ])، و روشهایی برای گنجاندن خودهمبستگی فضایی در مدلهای رگرسیون رایجتر شدهاند [ 25 ، 26 ،27 ].
Getis-Ord G i * یک آمار فضایی محلی برای اندازه گیری خودهمبستگی فضایی مثبت است که از Getis-Ord General G مشتق شده است (نگاه کنید به [ 28 ])، و محاسبه می کند که آیا یک مشاهده در یک خوشه از مقادیر زیاد یا پایین به عنوان تابعی از فاصله مقدار زیاد نشان میدهد که مقادیر بیشتر همسایگان آن نیز بالا هستند، که نشان میدهد که بخشی از یک نقطه داغ است، در حالی که مقدار پایین نشان میدهد که بیشتر همسایهها نیز پایین هستند و یک نقطه سرد را مشخص میکند [ 28 ] . این روش برای مطالعه نقاط داغ گونه های مختلف استفاده شده است [ 29 ، 30]، اما اخیراً مقیاس تحلیل به صراحت مورد توجه قرار گرفته است. مطالعات نشان داده اند که با افزایش اندازه محله، تعداد نقاط داغ شناسایی شده در منطقه مورد مطالعه نیز افزایش یافته است [ 31 ، 32 ]. با این حال، در مطالعهای بر روی توزیع علفهای هرز در استرالیا، لافان [ 33 ] دریافت که نقاط داغ با مقیاسهای متوسط مرتبط هستند و یک همسایگی وسیع منجر به حداکثر مقادیر G i * نمیشود. با این حال، تحقیقات کمی (در صورت وجود) برای بررسی دقیقتر الگوی فضایی پیچیده غنای گونهای انجام شده است.
ناایستایی فضایی رابطه یا فرآیندی را توصیف می کند که در سراسر فضا ثابت نیست [ 21 ]. اگر یک رابطه از نظر فضایی غیر ثابت باشد، آنگاه معیارهای آماری جهانی که برای بررسی آن استفاده میشوند، به احتمال زیاد نتایج مدل «صاف» را ایجاد میکنند که ممکن است فقط برای برخی از منطقه مورد مطالعه یا اصلاً قابل استفاده نباشد. غیر ایستایی روابط غنای گونه-محیط برای انواع گونه ها با استفاده از رگرسیون وزنی جغرافیایی (GWR) مورد مطالعه قرار گرفته است [ 15 ، 16 ، 17 ، 18 ، 19 ، 34 ].
GWR در مدلهای رگرسیون سنتی با محاسبه پارامترها (ضرایب، R2) برای هر مشاهده بر اساس مشاهدات همسایه پیشرفت میکند ، بنابراین به پارامترهای مدل اجازه میدهد تا در فضا متفاوت باشند. این مبنایی را برای آزمایش و بررسی غیرایستایی فضایی در رابطه بین پاسخ و متغیرهای پیشبینیکننده، و همچنین برای کشف چگونگی تأثیر این رابطه توسط مقیاس [ 35 ] فراهم میکند. مشابه آستانه فاصله مورد استفاده در G i *، همسایگی ها در GWR با استفاده از یک هسته فضایی تعریف شده توسط پهنای باند تعیین می شوند که می تواند ثابت (پهنای باند ثابت) یا تطبیقی (بر اساس تعداد مشاهدات) باشد.
آزمایش پهنای باند ثابت مختلف می تواند ابزار مفیدی برای کاوش در مقیاس فضایی و غیر ایستایی روابط عملکردی باشد. غذایی [ 15] تنوع فضایی در ضرایب GWR به دست آمده در چهار پهنای باند (1 درجه، 3 درجه، 5 درجه، 8 درجه) را برای رابطه بین غنای گونههای پرنده و بارش، دما و شاخص تفاوت عادی شده پوشش گیاهی (NDVI) در جنوب صحرای آفریقا شناسایی کرد. او نمونههایی از چهار رابطه غنای گونهای و بارندگی مختلف در منطقه مورد مطالعه بیان کرد: یک طرفه مثبت (ضریب در همه پهنای باند مثبت میماند)، یک طرفه منفی (منفی باقی میماند)، نسبتاً پایدار، و یکی که شیب علامت را از منفی تغییر میدهد. مثبت، با افزایش پهنای باند به 8 درجه، هر چهار به سمت تخمین جهانی همگرا می شوند. تنوع این روابط گونه-محیط نشان دهنده عدم ایستایی فضایی است (اگرچه به آستین [ 36 مراجعه کنید] برای بحث در مورد اینکه چگونه روابط منحنی مشخص شده نادرست ممکن است استنتاج نادرست از عدم ایستایی فضایی ایجاد کند، و جهت و نرخ تغییر در مشاهدات فردی که او در چهار پهنای باند ردیابی کرد، از نظر مکانی متغیر بود، که نشان میدهد مقیاس تأثیر مهمی بر روی دارد. این روابط
در سال های اخیر، رویکردهای سیستماتیک تری با استفاده از GWR برای مطالعه مقیاس روابط گونه-محیط ظاهر شده است [ 18 ، 34 ، 37 ]. آزبورن و همکاران در مطالعهای در رابطه بین گونههای لارک کالاندرا و زیست توده سبز، توسعههای شهری، ارتفاع و توپوگرافی در اسپانیا . [ 18 ] دریافتند که رابطه بین توزیع لارک و پیشبینیکنندههای محیطی فردی در مقیاسهای فضایی مختلف ثابت میشود، که نشان میدهد که توزیع تحت تأثیر فرآیندهایی است که در مقیاسهای چندگانه عمل میکنند. میلر و هانهام [ 34] دریافت که مقیاس روابط گونه-محیط برای گیاهان در صحرای موهاوه برای انواع مختلف گونهها (مثلاً نادر در مقابل معمول) و همچنین با انواع مختلف پیشبینیکنندههای محیطی (مانند آب و هوا در مقیاس وسیع در مقابل توپوگرافی پیچیده متفاوت است. ). ما و همکاران [ 37 ] عدم ایستایی غنای گونه های پرنده ایالت نیویورک را با استفاده از مدل پواسون فضایی در چهار تاخیر فضایی مورد مطالعه قرار داد و وارونگی هایی را در روابط غنا-محیط ( یعنی مثبت به منفی) و همچنین آمار پارامترهای محلی یافت. در مقیاس های مختلف این مطالعات اهمیت مطالعه سیستماتیک تعامل بین مقیاس و ناپایداری فضایی را تایید کردهاند.
مطالعات قبلی که تغییرات سیستماتیک در مقیاسها را برای آمارهای همبستگی مکانی محلی و غیرایستایی بررسی میکنند، مشخص کردهاند که تغییرات رخ میدهند، اما توزیع جغرافیایی این تغییرات را بررسی نکردهاند. جنبه جدایی ناپذیر آمار فضایی محلی «نقشهگذاری» آنها است و در حالی که مطالعات اخیر بر روی استانداردهای نقشهبرداری GWR با ارائه مقادیر ضرایب و مقادیر معنیداری به منظور کمک به تفسیر نتایج تمرکز کردهاند [ 38 ]، استفاده از نقشهها در شناسایی مکانهایی که ناپایداری رخ میدهد. (صرف نظر از اهمیت آماری) و اگر این تغییر در نتیجه پهنای باند باشد، یک ابزار اکتشافی قدرتمند خواهد بود و چیزی است که هنوز مورد استفاده قرار نگرفته است.
در اینجا ما روی دادههای غنای گونههای پرندگان با استفاده از اطلس پرندگان پرورشی نیویورک (BBA)، که توسط بیش از 1200 مشارکتکننده داوطلب در یک دوره پنج ساله جمعآوری شد، تمرکز میکنیم [ 39 ]. هدف از این تحقیق بررسی چگونگی تغییر مقیاس آمار فضایی محلی برای کشف عوامل محیطی مؤثر بر الگوهای غنای گونههای پرندگان در ایالت نیویورک است. این کار با استفاده از آمارهای خودهمبستگی و غیرایستایی جداگانه انجام شد. در مرحله اول، ما از آمار همبستگی خود برای کشف نقاط گرم و سرد غنای گونه ها استفاده می کنیم. ثانیاً، ما یک تغییر در رابطه غنای گونه-محیط زیست در سراسر منطقه مورد مطالعه را در نظر می گیریم که با تغییر قابل توجهی در مقدار ضریب نشان می دهد تا عدم ایستایی وابسته به مقیاس را نشان دهد.
2. مواد و روشها
2.1. غنای گونه ها و داده های زیست محیطی
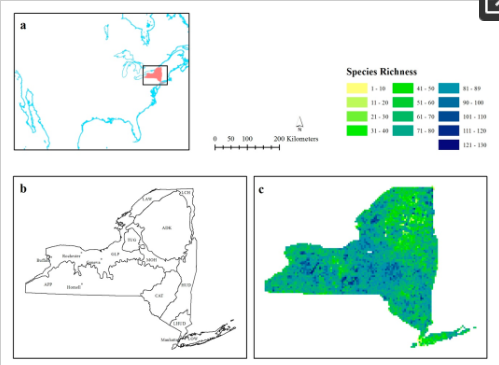
بررسی های BBA کل ایالت را در بلوک های بررسی 5 کیلومتر × 5 کیلومتر پوشش می دهد و شامل سه سطح رؤیت است. ممکن، محتمل و تایید شده است و غیبت ها تنها در صورتی ثبت می شوند که مجموعه ای از پروتکل های سختگیرانه رعایت شده باشد [ 40 ]. داده های اطلس جمع آوری شده از سال 2000 تا 2005 در این مطالعه مورد استفاده قرار گرفت [ 41 ]، با تنها مشاهده های تایید شده برای تولید غنای گونه ای ( شکل 1 ج). ایالت نیویورک از یازده اکوسون تشکیل شده است که از نظر وسعت فضایی متفاوت هستند ( شکل 1 ب)، و دارای عرض و طول تقریبی به ترتیب 450 کیلومتر و 500 کیلومتر است.
شکل 1. ( الف ) ایالت نیویورک در رابطه با آمریکای شمالی. ( ب ) منطقه مورد مطالعه با یازده اکوسون اصلی. مناطق بوم گردی عبارتند از، آدیرونداکس (ADK)، فلات آپالاچی (APP)، Catskills (CAT)، دشت های ساحلی (LOW)، دشت های دریاچه بزرگ، (GLP)، دره هادسون (HUD)، دریاچه شامپلین (LCH)، دره هادسون پایین ( LHUD)، دره Mohawk (MOH)، سنت لارنس (LAW)، و Tug Hill (TUG)؛ و ( ج ) داده های غنای گونه ای مورد استفاده در تجزیه و تحلیل.
بارش و میانگین حداقل دما از رگرسیون پارامتر-ارتفاع بر روی مدلهای شیب مستقل (PRISM)، ارتفاع در قالب مدل رقومی ارتفاع (DEM) از USGS و شاخص گیاهی تقویتشده (EVI) از طیف رادیومتر تصویربرداری متوسط (MODIS) محصول MOD13 [ 42 ] ( جدول 1 را ببینید ). EVI به جای پراکسی رایجتر مورد استفاده برای بهرهوری پوشش گیاهی، NDVI، استفاده شد، زیرا به تغییرات ساختار تاجپوش که به شدت بر مجموعههای پرندگان تأثیر میگذارد، پاسخگوتر است [ 43 ]. دادههای ماهانه برای همه متغیرها (به جز ارتفاع) از سال 2000 تا 2005 میانگینگیری شد. دادهها از وضوح اصلی خود نمونهبرداری شدند ( جدول 1 را ببینید).) با استفاده از تکنیک دوخطی تا وضوح 5 کیلومتر برای مطابقت با وضوح BBA. قبل از تجزیه و تحلیل، شش سلول شبکه، که غنای گونه ای صفر داشتند، حذف شدند. این شبکهها بر روی تودههای آبی گسترده (به عنوان مثال ، خلیج Peconic و دریاچه Oneida) قرار داشتند، و به همین دلیل از تجزیه و تحلیل به دلیل کمبود زمین که مانع از تکثیر پرندگان در آنجا میشد، حذف شدند. هر سلول شبکه BBA با کمتر از 50 درصد مقادیر EVI حاوی آب بیشتری نسبت به زمین در نظر گرفته شد و متعاقباً از تجزیه و تحلیل حذف شد ( n = 5332 اصلی، n = 5226 نهایی). تمام تجزیه و تحلیل در ArcMap 10.1 [ 44 ] انجام شد.
جدول 1. منبع و وضوح داده های مورد استفاده در تحقیق.
2.2. ناایستایی فضایی – تحلیل رگرسیون
کاوش در دادهها مشخص کرد که روابط بین غنای گونهها و متغیرهای محیطی خطی است (در راستای پیشنهاد آستین [ 36 ]). حداقل مربعات معمولی (OLS) و GWR متعاقباً برای بررسی روابط بین غنای گونههای پرنده و عوامل محیطی مورد استفاده قرار گرفتند. نسخه GWR مدل رگرسیون OLS را می توان به صورت زیر تعریف کرد:
در جایی که y متغیر وابسته است، مجموعه ای از m متغیر(های) مستقل وجود دارد، x k ، با k = 1، …، m ، و β 0i ( u ) نشان دهنده رابطه اطراف مکان u است.و مختص آن مکان است. یک ماتریس واریانس کوواریانس دارای وزن جغرافیایی برای تعریف هسته یا همسایگی استفاده شد که در آن مشاهدات استفاده شد. تأثیر همسایگان به صورت معکوس با فاصله از طریق یک طرح وزن دهی گاوسی محاسبه شد. پهنای باند یک اندازهگیری گاوسی از فاصله است که در همان واحدهای سیستم مختصات است و با بزرگتر شدن پهنای باند، وزنها به وحدت نزدیک میشوند و مدل GWR محلی به مدل OLS جهانی نزدیک میشود.
انتخاب پهنای باند هسته مناسب بسیار مهم است. پهنای باندی که خیلی کوچک است، یک مدل بیش از حد محلی ایجاد می کند و پهنای باند بیش از حد بزرگ، نتایجی نزدیک به مدل های جهانی ایجاد می کند. در نتیجه اکتشاف اولیه طیف وسیعی از پهنای باند (30 کیلومتر یا 6 سلول تا 450 کیلومتر یا 90 سلول)، ما تعیین کردیم که 120 کیلومتر فاصله ای است که در آن این روابط ثابت می شوند (جایی که پارامترهای مدل با افزایش پهنای باند دیگر تغییر نمی کنند) بنابراین ما روی چهار پهنای باند زیر تمرکز کردیم: 30 کیلومتر، 60 کیلومتر، 90 کیلومتر و 120 کیلومتر.
2.3. غنای گونه – تجزیه و تحلیل نقطه داغ و سرد
آماره G i * برای شناسایی خوشه های فضایی با غنای گونه ای زیاد یا کم استفاده شد . یک z – score مثبت نشان می دهد که مقادیر بیشتر همسایگان آن (محاسبه شده در متغیر فاصله) نیز بالا است، که نشان می دهد بخشی از یک نقطه داغ است، در حالی که یک z – score منفی نشان می دهد که همسایگان نیز پایین هستند. و یک نقطه سرد را شناسایی می کند [ 28 ]. فاصله را می توان تغییر داد، اندازه محله را تغییر داد، و بنابراین نقاط گرم و سرد را می توان به عنوان تابعی از مقیاس بررسی کرد. آستانه های فاصله مطابق با موارد مورد استفاده در تجزیه و تحلیل GWR برای بررسی اینکه چگونه اندازه گیری غنا وابسته به مقیاس بود استفاده شد. G i * ( d)آمار به صورت زیر تعریف می شود:
جایی که w ij عناصر ماتریس وزن ها هستند، i یک مکان خاص است، و d شعاع اطراف i است که برای تعریف همسایگان استفاده می شود، و n تعداد مکان ها است.
لازم به ذکر است که تعاریف همسایگی برای GWR و G i * به طور مستقیم قابل مقایسه نیستند. GWR از یک تابع وزن دهی با فاصله معکوس استفاده کرد، در حالی که G i * وزن یکسانی را برای همه همسایگان اعمال کرد. اما می توان آنها را از نظر بررسی تأثیر مقیاس بر الگوی فضایی مکمل یکدیگر دانست. همچنین متعاقباً اثرات لبه اجتنابناپذیری در امتداد مرز ایالت نیویورک وجود خواهد داشت، زیرا مرزهای ایالتهای زمینی لزوماً پایان وسعت نیستند، و در حالی که محدودیتهای داده ما را از گنجاندن مشاهدات فراتر از این مرزها باز میدارد، این چیزی است که باید به آن توجه شود.
3. نتایج
هر چهار متغیر استاندارد شده در مدل نهایی OLS معنی دار بودند ( جدول 2 ). بارش و ارتفاع رابطه منفی با غنای گونه ای پرندگان داشتند، احتمالاً به دلیل تحمل فیزیولوژیکی معکوس مرتبط با بارندگی بیشتر در نزدیکی ساحل و دماهای پایین تر در ارتفاعات بالاتر، در حالی که EVI و دمای حداقل رابطه مثبتی با مزایای پوشش گیاهی بیشتر داشتند. لانه سازی و اهمیت شرایط گرمتر در بقای بسیاری از پرندگان دلیل این روابط است. متغیری که بیشترین تأثیر را در مدل نهایی داشت EVI بود.
جدول 2 همچنین آمار توصیفی مدل های نهایی GWR را در چهار مقیاس آزمایش شده در اینجا نشان می دهد. بارش که رابطه منفی با غنای گونه ای در مقیاس جهانی داشت، برای هر چهار پهنای باند یک رابطه مثبت متوسط داشت. به طور مشابه، با وجود رابطه مثبت جهانی، دما در پهنای باندهای کوچکتر یک رابطه میانگین منفی داشت. هر چهار متغیر محیطی دارای یک محدوده ضریب بودند که به موجب آن حداقل مقدار منفی و حداکثر مقدار مثبت بود ( جدول 2 ). ارتفاع مهمترین متغیر در هر چهار پهنای باند بررسی شده در تحلیل GWR بود، اما از نظر اهمیت در مدل نهایی OLS سومین متغیر بود.
جدول 2. نتایج رگرسیون از حداقل مربعات معمولی (OLS) و رگرسیون وزنی جغرافیایی (GWR) (تست شده در چهار پهنای باند). * معنی دار در 0.05، ** معنی دار در 0.01.
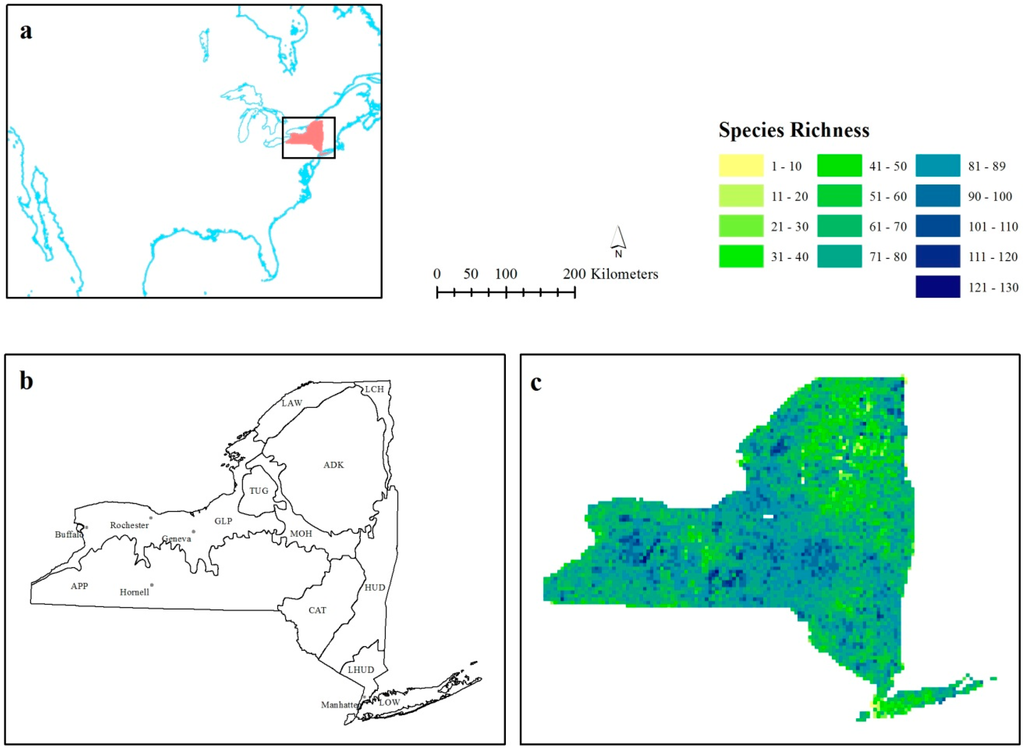
شکل 2. مقادیر R2 محلی برای هر مشاهده در مقایسه با حداقل مربعات معمولی جهانی (OLS) R2 در پهنای باند ( a ) 30 کیلومتر، ( b ) 60 کیلومتر، ( c ) 90 کیلومتر، و ( d ) 120 کیلومتر.
میانگین مقدار R2 تنظیم شده در هر چهار پهنای باند در مقایسه با مدل OLS ( جدول 2 ) بالاتر بود ، با این حال، حداقل مقدار R2 محلی نشان می دهد که برخی از مشاهدات R2 کمتری نسبت به آمار OLS جهانی داشتند. در حالی که این تعجب آور نیست (مدل OLS جهانی عملاً یک GWR است که در آن همه مکانها وزن برابر دارند)، مطالعات GWR اغلب فقط بر مقادیر R2 محلی متمرکز هستند که بالاتر از مدل OLS جهانی هستند و توجه کمی به توزیع فضایی داده میشود. این ارزش ها در پهنای باند 30 کیلومتری ( شکل 2 الف)، 12 “خوشه” به هم پیوسته فضایی با مقادیر پایین تر R2 وجود دارد، و با افزایش پهنای باند به 90 کیلومتر (شکل 2 ج)، فرکانس و وسعت آنها کاهش می یابد. در 90 کیلومتری، اکثر مشاهدات R2 پایین تر در موقعیت جغرافیایی خود ثابت به نظر می رسند.
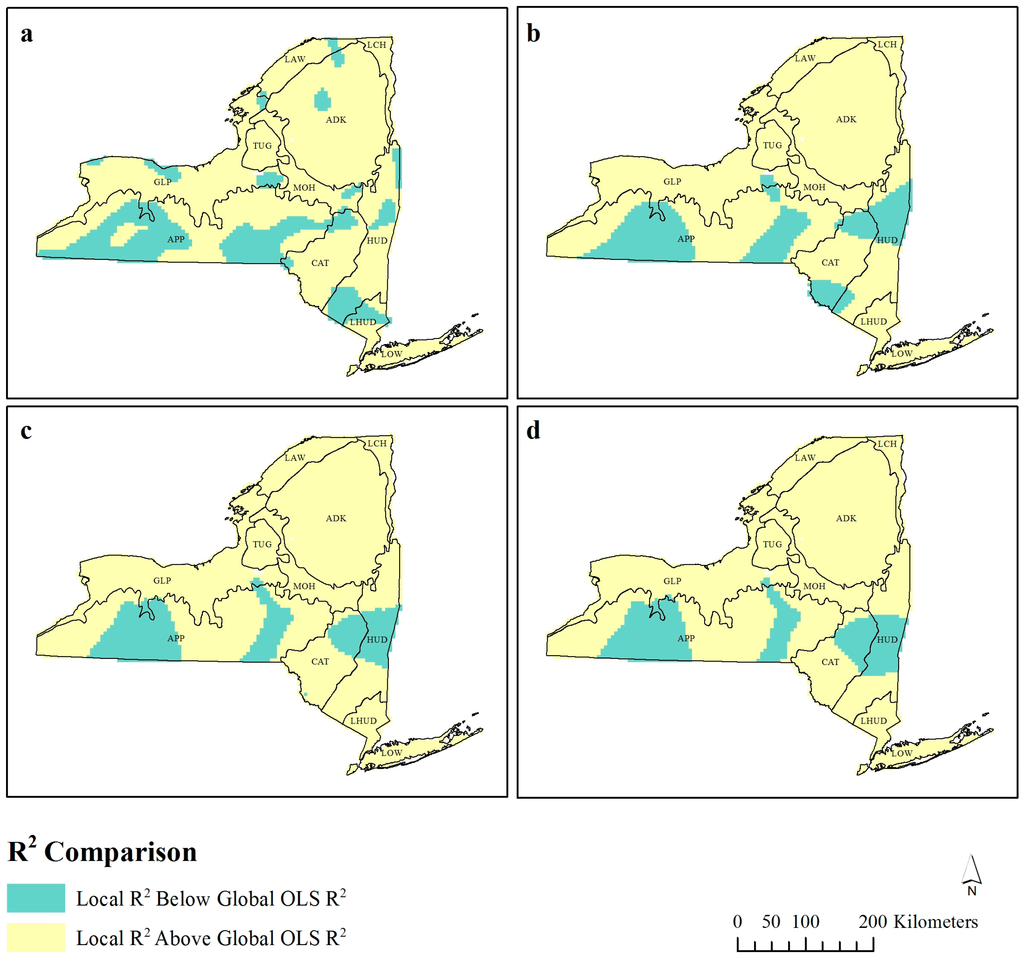
شکل 3. متغیر استاندارد شده که بیشترین اهمیت را برای پهنای باند رگرسیون وزنی جغرافیایی (GWR) دارد ( a ) 30 کیلومتر، ( ب ) 60 کیلومتر، ( c ) 90 کیلومتر، و ( d ) 120 کیلومتر. فقط متغیرهایی با β > 1.96 گزارش شده است.
این کاوش فضایی مقادیر R 2 زمانی مهم است که شکل 2 با شکل 3 و شکل 4 مقایسه شود، زیرا الگوها شروع به ظهور می کنند. نواحی مقادیر R2 محلی پایین تر ( شکل 2 ) در کنار تغییرات در ضریب تسلط استاندارد شده قابل توجه (96/1±β) قرار دارند ( شکل 3 ). ناحیه کوهاندار با مقادیر پایینتر R2 که در هر چهار پهنای باند در غرب ایالت و APP اکوزون گزارش شده است ( شکل 2 ) در امتداد مرزی قرار دارد که EVI و دما ضرایب غالب هستند ( شکل 3 ). به همین ترتیب، مناطق جمع شده R پایین تر2 مقدار در شرق ایالت ( شکل 2 ) مربوط به مرزی است که در آن بارش یا دما مهم ترین در نظر گرفته می شود ( شکل 3 b-d).
به طور مشابه، ناحیه برآمدگی مقادیر R2 محلی پایین تر ( شکل 2 ) با ناحیه کوهان دار در شکل 4 c مطابقت دارد که ضریب وارونگی دما را نشان می دهد. به نظر می رسد که نوارهای ضرایب مثبت با افزایش پهنای باند به ضرایب منفی تغییر می کنند (DEM: داده نشان داده نشده است). همانطور که پهنای باند افزایش می یابد، دره بعدی در سراسر رابطه خود را با دما و غنا معکوس می کند، که به احتمال زیاد نتیجه افزایش تغییرات غنای همسایگی بین کف دره های مولد و خط الرأس های کم بازده است. در واقع، به نظر میرسد وارونگیهایی برای همه متغیرهای مستقل ( شکل 4 ) در هر سه ناحیه با مقادیر پایینتر R2 وجود دارد .شکل 2 ).
شکل 4. مشاهداتی که روابط غنا-محیط 30 کیلومتری خود را با افزایش مقیاس برای ( الف ) بارش، ( ب ) شاخص پوشش گیاهی افزایش یافته (EVI)، ( ج ) دما، و ( d ) ارتفاع معکوس میکنند.
شکل 4 فاصله پهنای باندی را نشان می دهد که در آن ضرایب رابطه اساسی خود را با غنای گونه ای معکوس می کنند ( یعنی از مثبت به منفی یا بالعکس). بارش دارای سه منطقه مجزا از روابط مثبت و منفی است که هر یک از این مناطق با افزایش پهنای باند گسترش می یابند. در بخش شمال شرقی ایالت، ناحیه رابطه غنای منفی بارندگی محدود است و سه نوار تیرهتر قرمز را میتوان برای نشان دادن این موضوع مشاهده کرد. به نظر می رسد که دما در پهنای باند مختلف روابط مختلفی دارد، اگرچه در مقایسه با سایر متغیرها، مشاهدات کمتری وجود دارد که رابطه اساسی آنها را با غنای گونه ای معکوس نمی کند. به طور خاص، در جنوب غربی ایالت در فلات آپالاچی (اپزون APP)، یک شکل کوهان دار وجود دارد، جایی که در 30 کیلومتری کل منطقه مشاهدات مثبت دارد و با افزایش پهنای باند، نیمی از این منطقه به یک رابطه منفی تغییر می کند. . در 120 کیلومتری اکثر منطقه مورد مطالعه دارای یک رابطه غنای-EVI مثبت است، به جز منطقه در ADK اکوزون. مناطق اطراف منطقه روابط منفی EVI در 120 کیلومتر در پهنای باند کمتر منفی بودند و متعاقباً با افزایش پهنای باند، ناحیه رابطه منفی مثبت می شود (مانند ضریب OLS جهانی). ارتفاع یک رابطه عمدتاً منفی در سراسر ایالت دارد، به جز نوار در وسط، که یک رابطه مثبت دارد. بسیاری از مشاهدات منفی معکوس نمی شوند، با میزان زیادی از مشاهدات مداوم که در هیچ فاصله ای تغییر نمی کنند. تعداد انگشت شماری مشاهدات در لبه ناحیه مثبت وجود دارد که با افزایش پهنای باند از منفی به مثبت تغییر می کند و برخی در قسمت جنوب غربی ایالت. مناطق اطراف منطقه روابط منفی EVI در 120 کیلومتر در پهنای باند کمتر منفی بودند و متعاقباً با افزایش پهنای باند، ناحیه رابطه منفی مثبت می شود (مانند ضریب OLS جهانی). ارتفاع یک رابطه عمدتاً منفی در سراسر ایالت دارد، به جز نوار در وسط، که یک رابطه مثبت دارد. بسیاری از مشاهدات منفی معکوس نمی شوند، با میزان زیادی از مشاهدات مداوم که در هیچ فاصله ای تغییر نمی کنند. تعداد انگشت شماری مشاهدات در لبه ناحیه مثبت وجود دارد که با افزایش پهنای باند از منفی به مثبت تغییر می کند و برخی در قسمت جنوب غربی ایالت. مناطق اطراف منطقه روابط منفی EVI در 120 کیلومتر در پهنای باند کمتر منفی بودند و متعاقباً با افزایش پهنای باند، ناحیه رابطه منفی مثبت می شود (مانند ضریب OLS جهانی). ارتفاع یک رابطه عمدتاً منفی در سراسر ایالت دارد، به جز نوار در وسط، که یک رابطه مثبت دارد. بسیاری از مشاهدات منفی معکوس نمی شوند، با میزان زیادی از مشاهدات مداوم که در هیچ فاصله ای تغییر نمی کنند. تعداد انگشت شماری مشاهدات در لبه ناحیه مثبت وجود دارد که با افزایش پهنای باند از منفی به مثبت تغییر می کند و برخی در قسمت جنوب غربی ایالت. و متعاقباً با افزایش پهنای باند، ناحیه رابطه منفی مثبت می شود (مانند ضریب OLS جهانی). ارتفاع یک رابطه عمدتاً منفی در سراسر ایالت دارد، به جز نوار در وسط، که یک رابطه مثبت دارد. بسیاری از مشاهدات منفی معکوس نمی شوند، با میزان زیادی از مشاهدات مداوم که در هیچ فاصله ای تغییر نمی کنند. تعداد انگشت شماری مشاهدات در لبه ناحیه مثبت وجود دارد که با افزایش پهنای باند از منفی به مثبت تغییر می کند و برخی در قسمت جنوب غربی ایالت. و متعاقباً با افزایش پهنای باند، ناحیه رابطه منفی مثبت می شود (مانند ضریب OLS جهانی). ارتفاع یک رابطه عمدتاً منفی در سراسر ایالت دارد، به جز نوار در وسط، که یک رابطه مثبت دارد. بسیاری از مشاهدات منفی معکوس نمی شوند، با میزان زیادی از مشاهدات مداوم که در هیچ فاصله ای تغییر نمی کنند. تعداد انگشت شماری مشاهدات در لبه ناحیه مثبت وجود دارد که با افزایش پهنای باند از منفی به مثبت تغییر می کند و برخی در قسمت جنوب غربی ایالت. بسیاری از مشاهدات منفی معکوس نمی شوند، با میزان زیادی از مشاهدات مداوم که در هیچ فاصله ای تغییر نمی کنند. تعداد انگشت شماری مشاهدات در لبه ناحیه مثبت وجود دارد که با افزایش پهنای باند از منفی به مثبت تغییر می کند و برخی در قسمت جنوب غربی ایالت. بسیاری از مشاهدات منفی معکوس نمی شوند، با میزان زیادی از مشاهدات مداوم که در هیچ فاصله ای تغییر نمی کنند. تعداد انگشت شماری مشاهدات در لبه ناحیه مثبت وجود دارد که با افزایش پهنای باند از منفی به مثبت تغییر می کند و برخی در قسمت جنوب غربی ایالت.
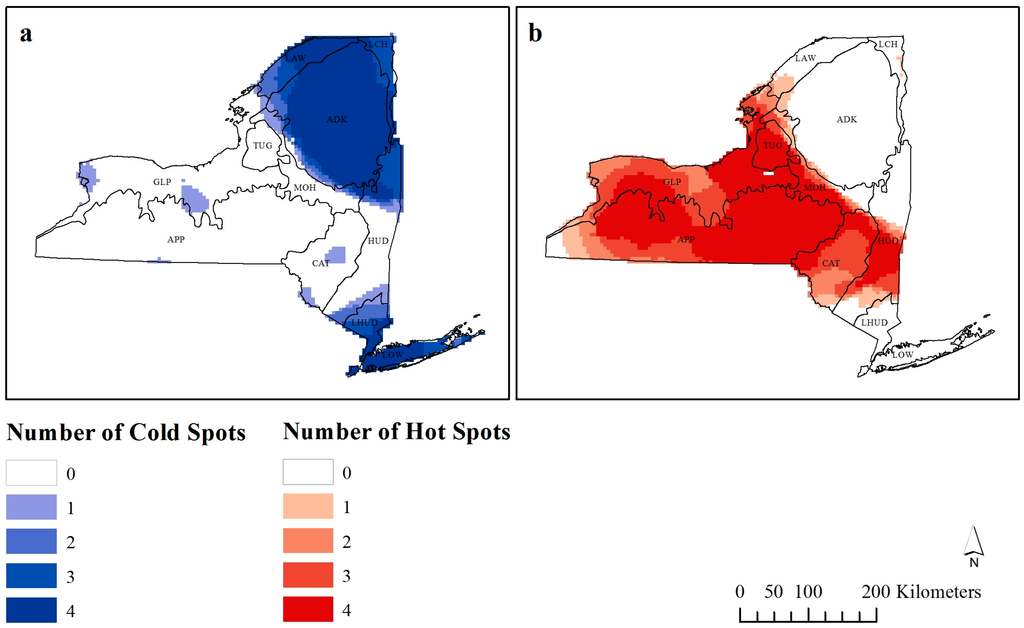
شکل 5. تعداد دفعاتی که هر مشاهده توسط تحلیل Getis-Ord G i * یک ( a ) نقطه سرد یا ( b ) نقطه داغ در نظر گرفته می شود.
از 5226 مشاهدات، با افزایش فاصله آستانه فاصله آماره G i * به 120 کیلومتر، 3173 مشاهده نقطه داغ و 2053 مشاهده نقطه سرد در نظر گرفته شد. شکل 5 نشان می دهد که چند بار هر مشاهده به عنوان نقطه گرم یا سرد قابل توجه ( z – score > 1.96) در پهنای باند مختلف در نظر گرفته می شود. رنگهای تیرهتر بهعنوان نقاط سرد یا گرم مستقل در مقیاس در نظر گرفته میشوند، زیرا در پهنای باند بیشتری به این صورت طبقهبندی میشوند.
4. بحث
یک نتیجه ثابت از مطالعات مقایسه GWR افزایش GWR محلی R2 در مقایسه با OLS جهانی R2 است [ 15 ، 16 ، 18 ، 45 ]. به عنوان مثال، فودی [ 15 ] دریافت که بیش از 90 درصد از تنوع بین غنای گونههای پرندگان جنوب صحرای آفریقا و مجموعهای از متغیرهای محیطی (اقلیم و NDVI) در مقیاسهای خوب (1 درجه) توضیح داده شده است، و واریانس توضیحدادهشده کاهش مییابد. با افزایش پهنای باند مورد استفاده در GWR (به 8 درجه). از جدول 2 می توان مشاهده کرد که میانگین مقدار R 2 محلی در همه پهنای باند بالاتر از OLS R 2 جهانی است.، از این یافته های قبلی پشتیبانی می کند.
مقادیر R2 محلی تولید شده با GWR عموماً بالاتر از R2 جهانی است به دلایلی که بیشتر به برازش مدل محلی مربوط می شود تا افزایش واقعی واریانس توضیح داده شده [ 34 ، 46 ]. در حالی که R2 محلی ممکن است قوی ترین آمار محاسبه شده توسط GWR برای بررسی غیرایستایی نباشد، هنوز مشاهداتی وجود دارد که R2 محلی کمتری نسبت به مقدار OLS R2 جهانی دارند و شکل 2 مشخص می کند که این مشاهدات در کجای ایالت نیویورک قرار دارند. ، با همگن شدن الگوی فضایی این مشاهدات در 90 کیلومتری.
شباهت در مناطقی که مقدار R2 محلی کمتری نسبت به مقدار جهانی OLS نشان میدهند ( شکل 2 ) و مناطقی که وارونگی را در رابطه غنا-محیط نشان میدهند ( شکل 4 ) نشان میدهد که مناطق با مقدار R2 محلی پایینتر شاید در رابطه غنا-محیط بیشترین ناپایداری است و مدلهای نهایی احتمالاً واریانس را به دلیل عوامل دیگری که در مدلهای ما لحاظ نشده است توضیح میدهند. بنابراین، هر گونه رابطه غنا-محیط مشاهده شده در این مناطق باید با احتیاط رفتار شود، زیرا این رابطه ثابت نیست و مشاهدات R2 پایین است. با این حال، این فرض نمیکند که مکانهایی با R محلی بیشتر2 نسبت به OLS R2 جهانی ایستایی را نشان داد، زیرا چندین رابطه غیر ثابت در این مکان ها مشاهده شد، به ویژه دما و بارش ( شکل 4 a,c).
Ecozone ADK که شامل پارک آدیرونداک (بزرگترین منطقه حفاظت شده در سطح ایالت در ایالات متحده آمریکا) است، یک رابطه مستقل از مقیاس با EVI را نشان داد ( شکل 4 ). در حالی که رابطه معکوس بین EVI و غنای گونه های پرندگان ممکن است تعجب آور باشد، این رابطه غیر معمول نیست. در حالی که پارک آدیرونداک دارای جمعیت ثابتی از پرندگان است، بسیاری از این پرندگان گونههای خاص جنگل هستند و بنابراین غنای گونهای بهویژه زیاد نخواهد بود. تکه تکه شدن مناطق اطراف ممکن است فراوانی یک گونه خاص را به دلیل از دست دادن زیستگاه کاهش دهد، اما زیستگاه های متعددی را در این منطقه ایجاد می کند که به پرندگانی که معمولاً با این زیستگاه مرتبط نیستند در اطراف لبه ها یافت می شوند [ 47 ].]، در نتیجه غنای گونه ای افزایش می یابد. بنابراین، افزایش EVI برای مناظر تکه تکه شده، غنا را افزایش می دهد، اما در منظر جنگل پیوسته اینطور نیست. این روش تجسم نتایج، رابطه غنا-EVI را در زمینه جغرافیایی قرار می دهد و تفسیر را معنادارتر می کند.
ایستایی در رابطه غنا-محیط که اکوسون دشت های ساحلی (LOW) در هر چهار رابطه غنا-محیط نشان داد ( شکل 4 ) را می توان به منحصر به فرد بودن این ناحیه نسبت داد. ارتفاع پایین تر و نزدیکی نزدیک به آب هر دو بر نتایج مشاهده شده در اینجا تأثیر می گذارد، زیرا این شرایط در بقیه ایالت دیده نمی شود و منجر به الگوهای غنای گونه ای می شود که در جاهای دیگر دیده نمی شود. ما مشاهداتی که بیش از 50 درصد آب داشتند را حذف کردیم تا برخی از این تأثیرات را نفی کنیم و همچنین هرگونه سوگیری را که ممکن است از طریق تأثیر منطقه گونه مشاهده شود حذف کنیم (نگاه کنید به [ 48 ]] برای بحث در مورد رابطه گونه-منطقه). پرندگان دریایی در محاسبات غنای گونه ای گنجانده شدند و در حالی که تفاوت بین پرندگان دریایی و خشکی ممکن است به روابط محیطی متفاوتی منجر شود، تمایز بین انواع مختلف گونه های پرندگان منجر به نتایج غیرقابل کنترلی می شد و اقدامات احتیاطی برای حذف مشاهدات انجام شد. برای کاهش هرگونه سوگیری احتمالی ناشی از صفات فیزیولوژیکی مختلف در پرندگان کافی است. گستردگی منهتن و دیگر مراکز شهری نیز ممکن است تأثیر منفی زیادی بر غنای منطقه داشته باشد. متعاقباً تعدادی فرآیند مستقل از مقیاس در اکوزون LOW وجود دارد.
زمانی که تأثیر منهتن و همچنین تأثیرات محیطی ساحلی خاص در بالای جزیره در نظر گرفته میشود، یافتن این موضوع که منطقه اطراف شهر نیویورک به طور مداوم یک نقطه سرد است، تعجب آور نبود. همچنین یک نقطه سرد در شمال شرقی ایالت وجود دارد، جایی که پارک آدیرونداک در آن یافت می شود، که دلایل این غنای کمتر قبلاً مورد بحث قرار گرفته است. مناطق غنی مثبت واقعی شامل منطقه بین بوفالو، روچستر و هورنل (موقعیت های ارائه شده در شکل 1 ب) و همچنین منطقه در سراسر مرکز ایالت بود.
نکته قابل توجه باید شش ناحیه ای باشد که هر دو نقطه گرم و سرد در برخی از پهنای باند هستند ( شکل 5 ). پنج مورد از اینها در خوشه نقاط داغ بزرگتر یافت می شوند، که نشان می دهد این مناطق دارای نقاط سرد در مقیاس محلی (یا غنای کم) هستند. این مناطق شامل بوفالو و ژنو (و مناطق اطراف آنها) است. در حالی که اخیراً تغییری به سمت افزایش غنای گونههای پرندگان در مناطق شهری به دلیل منابع غذایی تأمینشده توسط انسان بهویژه در فصل زمستان صورت گرفته است [ 49 ]]، به نظر نمی رسد نیویورک از این روند پیروی کند. در حالی که هیچ دلیل واضحی در پشت این نقاط سرد محلی وجود ندارد، این روش تجسم به مناطقی اشاره می کند که به تحقیقات بیشتری نیاز دارند که چرا این مناطق دارای غنای کم در مقیاس های محلی هستند اما در مقیاس های وسیع تر، نقاط داغ محسوب می شوند.
مطالعاتی که تغییرات در نتایج G i * را بررسی کردهاند، به طور کلی افزایش نقاط داغ را در مقیاسهای وسیعتر، بدون توجه به پدیدهها نشان دادهاند [ 31 ، 32 ]، اگرچه مقیاسهای میانی منجر به تعداد بهینه نقاط داغ میشوند. [ 33 ] را ببینید). نتایج ما با این موافق است، و بنابراین، با افزایش مقیاس تحقیق، دادهها به احتمال زیاد به عنوان نقاط داغ خوشهبندی میشوند. لازم به ذکر است که نلسون و بوتس [ 32 ] و دی کاسترو و همکاران. [ 31 ] تشخیص نقاط گرم و سرد را به ترتیب بر روی میزان آلودگی سوسک کاج کوهی و میزان آلودگی مالاریا مطالعه کردند، در حالی که Laffan [ 33] تراکم علف های هرز را مورد مطالعه قرار داد. مطالعه ما از غنای گونه ای استفاده کرد و متعاقباً به طور مستقیم با این مطالعات دیگر قابل مقایسه نیست. بنابراین، اگر قرار است از این تحلیل فضایی در کاربردهای اکولوژیکی استفاده شود، تحقیقات بیشتری در مورد چگونگی تأثیر مقیاس بر تشخیص نقاط گرم و سرد غنای گونهای باید انجام شود.
ویتاکر [ 50] تعدادی از راه هایی را که در آنها می توان تنوع گونه ها را در نظر گرفت توضیح داد. تنوع آلفا به تعداد کل گونه ها در یک زیستگاه اشاره دارد و در مطالعه ما از طریق استفاده از غنای گونه ای در نظر گرفته شد. تنوع بتا، تمایز گونه ها بین دو زیستگاه متمایز، و همچنین تنوع گاما، ترکیبی از تنوع آلفا و بتا نیز می تواند مورد استفاده قرار گیرد. در حالی که غنای گونهای برای شناسایی تنوع زیستی یک منطقه بسیار مفید است، نمیتوان از آن برای تعیین اینکه آیا مجموعهای از مشاهدات مجاور با غنای بالا حاوی مجموعههای یکسانی از گونهها یا کاملاً متفاوت است، استفاده کرد. کار آینده باید به بررسی تأثیر مقیاس و آمار فضایی محلی برای اندازه گیری تنوع بتا و گاما هدایت شود.
5. نتیجه گیری ها
شناسایی غنای گونهها و روابط محیطی، و گنجاندن روشهایی برای شناسایی و تحلیل همبستگی فضایی و غیرایستایی عناصر حیاتی برای حفاظت هستند. استفاده از آمارهای مکانی محلی میتواند راهی مستقیم برای ارزیابی و تجسم روابط غنا-محیط فراهم کند. در حالی که مطالعات به چگونگی تأثیر تغییرات در مقیاس بر نتایج بهدستآمده از آمارهای فضایی محلی نگاه کردهاند، هنوز یک حوزه تحقیقاتی نسبتاً مورد مطالعه قرار نگرفته است، و با تغییر سیستماتیک اندازههای محله، میتوان تشخیص داد که این روابط در مقیاسهای چندگانه چقدر پایدار هستند. ما دریافتیم که تفاوت های محلی قابل توجهی در روابط بین EVI، بارش، ارتفاع، دما و غنای گونه های پرندگان در تعاریف مختلف محله ها وجود دارد. که پیامدهای مهمی برای هر استراتژی مدیریتی اعمال شده بر روی نتایج چنین تحلیلی دارد. این تغییرات در کجا رخ می دهند و در چه پهنای باندی، جنبه های مهم تحلیل اکتشافی هستند و در حالی که مطالعات قبلی (به عنوان مثال، [15 ) روابط گونه-محیط را برای مشاهدات فردی “ردیابی” کرده اند، زیرا مقیاس آمار فضایی محلی به طور سیستماتیک تغییر می کند، با توسعه یک تکنیک تجسم جدید برای ترسیم این تغییرات مقیاس در آمار فضایی محلی، مکان غیر ایستایی و خود همبستگی می تواند در زمینه جغرافیایی، در کل منطقه مورد مطالعه تفسیر شود. وارونگی در رابطه غنا-محیط با مقیاس (مثلاً از مثبت به منفی) می تواند پیامدهای جدی بر هر استراتژی مدیریتی مبتنی بر تجزیه و تحلیل داشته باشد، بنابراین محققان باید این الگوها را در مقیاس های مختلف مطالعه کنند. این تکنیک تجسم به ما این امکان را داد تا الگوهایی را شناسایی کنیم که شروع به ظهور بین غیرایستایی ، R2 محلی کردندارزش ها و متغیرهایی که بیشترین اهمیت را دارند. استفاده از آمارهای مکانی محلی امکان تجزیه و تحلیل محیطی دقیقتر را فراهم میکند و فرصتی را برای کشف الگوهای غنای گونهها در مقیاسهای فضایی مختلف در تجسمهای مختلف ارائه میدهد.




بدون نظر