1. مقدمه
درک واکنش اکولوژیکی به تغییرات محیطی انسانی، از جمله تغییرات آب و هوا، کاربری زمین، پوشش زمین و سایر عوامل، نیازمند ابزارهای کمی برای توصیف، تحلیل و تجسم تغییرات پویا در جمعیت ارگانیسمهای کلیدی مورد علاقه است. توسعه چنین ابزارهایی با این واقعیت دشوار می شود که پاسخ های جمعیتی به تغییرات محیطی از نظر مکانی و زمانی پیچیده است، به ویژه برای موجودات با مراحل زندگی محیطی متعدد، مانند آنهایی که در انتقال بیماری های منتقله از طریق ناقل (VBD) شرکت می کنند. جمعیتهای ناقل بیماری ممکن است در زمانبندی و مدت زمان تغییرات فصلی را نشان دهند و پاسخ عموماً غیرخطی آنها به سیگنالهای محیطی، پیشبینی خطر ناشی از VBD را تحت شرایط محیطی تغییریافته چالشبرانگیز میکند [ 1 ]، 2 ، 3 ]. هنگامی که در معرض تغییر شرایط آب و هوایی قرار می گیرد، توزیع برداری و خطرات VBD ممکن است به طور قابل ملاحظه ای در طول زمان و مکان تغییر کند [ 1 ، 4 ، 5 ]. با این حال، مقدار زیادی عدم قطعیت برای بسیاری از سیستم های VBD [ 4 ، 6 ، 7 ، 8 ] باقی مانده است، و اطلاعات کمی در مورد ماهیت پویای واکنش جمعیت به تغییرات آب و هوایی، به ویژه فنولوژی برداری (زمان بندی مراحل زندگی)، فصلی و مدت زمان رویدادهای کلیدی جمعیت
در حالی که برخی از تحلیلهای اکولوژیکی پاسخ جمعیتی پویای گونههای مختلف گیاهی و بندپایان را به فشارهای بیرونی به شکلی واضح مشخص کردهاند (به عنوان مثال، [ 9 ، 10 ، 11 ، 12 ])، بسیاری از کارهای مشابه روی VBD، حوزه فضایی را نادیده گرفته است [ 13 ]. ، 14 ]. با این حال، کارهای دیگر از پویایی سیستم چشم پوشی می کنند، به جای آن، الگوهای فضایی معیارهای جمعیت ساکن، مانند حضور/غیاب یا فراوانی میانگین را بررسی می کنند (به عنوان مثال، [ 15 ] برای بیماری لایم و [ 16 ] را ببینید.] برای هانتاویروس). چنین تجزیه و تحلیل هایی از روابط آماری بین آب و هوا و تناسب زیستگاه برای تخمین، به عنوان مثال، تغییرات بالقوه در توزیع مناسب زیستگاه برای، یا تراکم پوره، Ixodes scapularis ، ناقل بیماری لایم [ 17 ، 18 ] استفاده می کند. این رویکرد بینش کمی در مورد ماهیت واکنش جمعیت در طول زمان ارائه میکند، مانند تغییرات در زمان اوج جمعیت یا تغییر در تراکم جمعیت در طول دورههای قرار گرفتن در معرض کلیدی (به عنوان مثال، فصل بالا برای فعالیتهای تفریحی). با توجه به اختلاف نظر اساسی و مداوم در مورد اینکه چگونه آب و هوا ممکن است توزیع VBD را تغییر دهد (به عنوان مثال، [ 4 ، 7])، تحلیل هایی که قادر به ارزیابی رابطه بین اجبارهای برون زا و پویایی جمعیت در فضا و زمان هستند، ممکن است چنین بینشی را ارائه دهند.
علاوه بر این، تجسم جغرافیایی پاسخ پویا VBD به تغییرات محیطی میتواند اطلاعات کلیدی (به عنوان مثال، نقشههایی که پدیدههای پیچیده مکانی-زمانی را خلاصه میکنند) برای توسعه سیاستهایی برای پاسخ به ریسک در حال تغییر فراهم کند. بنابراین، ابزارهای مکانی برای توصیف، تجزیه و تحلیل و پیشبینی پاسخ VBD به تغییرات آینده مطلوب هستند، و اینها باید بر پدیدههای دینامیکی که برای درک ریسک مهم هستند، مانند فنولوژی برداری و فصلی تأکید کنند. فنولوژی – زمان مراحل زندگی – به عنوان حساس به تغییرات آب و هوایی شناخته شده است و یک عامل تعیین کننده مهم در توزیع فضایی بندپایان است [ 19 ، 20 ]]. مدلهای فعلی که توزیع بندپایان را در اقلیمهای آینده بررسی میکنند، عموماً فنولوژی را نادیده میگیرند، در عوض، رابطهای بین فراوانی فعلی یک بردار و ویژگیهای کلیدی زیستگاه برقرار میکنند و سپس، آن مدل را برای شرایط آینده پیشبینیشده به کار میبرند [ 21 ، 22 ]. بررسی واکنش فنولوژیکی یک موجود زنده می تواند تأثیرات مهم، اما ظریف تغییر آب و هوا را نشان دهد. به عنوان مثال، تاریخ گلدهی و باردهی تعیینکنندههای مهمی در توزیع آسپن هستند [ 23 ]، و تاریخ اولین تخمگذاری برای توزیع شب پره کولی مهم است [ 24 ].]. مشخص کردن پاسخهای دینامیکی خاص مرحله زندگی میتواند چنین عوامل تعیینکننده ظریفی را در توزیع بردارها در آب و هوای آینده برجسته کند.
فصلی بودن رویدادها نیز ممکن است تحت شرایط آینده تغییر کند و پیامدهای مهمی برای خطر VBD داشته باشد. برای مثال، جمعیت های ناقل ممکن است در زمان های خاصی از سال به اوج خود برسند، با اوج بروز بیماری در زمان های دیگر (به عنوان مثال، [ 25 ] برای بیماری لایم را ببینید). برخی از مدلهای پاسخ VBD به تغییرات آب و هوایی تلاش میکنند تغییرات فصلی را تقریباً مشخص کنند (به عنوان مثال، [ 26 ]). برخی عناصر فصلی، مانند دما، رطوبت و ساعات روز را از طریق مدلهای درجه روز (مثلاً [ 27 ، 28 ، 29 ، 30 ]) ادغام میکنند، و هنوز، برخی دیگر به صراحت فصلی بودن را در نظر نمیگیرند (به عنوان مثال، [ 31 ]]). یک نمایش فضایی دقیق تر از تغییرات فصلی، توصیف تأثیر بالقوه عمیقی که تغییر محیطی ممکن است بر طول و زمان فصول انتقال VBD داشته باشد را ممکن می سازد.
در اینجا، ما یک رویکرد مدلسازی صریح فضایی را برای بررسی پاسخهای جمعیتی پویا یک ناقل بیماری مورد علاقه، با هدف افزایش درک خود از خطر VBD آینده، توسعه میدهیم. ما مفهوم ویژگیهای جمعیت پویا (DPF) را معرفی میکنیم، که اطلاعاتی در مورد چرخه جمعیت، زمانبندی فصلی و رویدادهای فنولوژیکی در مراحل زندگی بردار ارائه میدهد. مهمتر از همه، ما توضیح می دهیم که چگونه تجزیه و تحلیل چنین ویژگی هایی – مانند تعداد و زمان اوج جمعیت ( جدول 1 ) – ممکن است برای پیش بینی خطر بیماری استفاده شود.
برای نشان دادن کاربرد این رویکرد مدلسازی، ما پاسخهای کنه گوزن پا سیاه ( Ixodes scapularis )، ناقل بیماری لایم را به تغییرات دما در سراسر شرق ایالات متحده بررسی میکنیم. I. scapularis یک ارگانیسم مدل عالی است که با آن می توان تأثیر تغییرات آب و هوا بر ویژگی های فنولوژیکی و فصلی را بررسی کرد: شناخته شده است که به شرایط محیطی از جمله دما بسیار حساس است [ 6 ، 15 ، 25 ، 32 ]. علاوه بر این، سه مرحله زندگی I. scapularis (لارو، پوره و بالغ) به شرایط دمایی متفاوتی برای حمایت از یافتن میزبان یا پیشرفت به مرحله بعدی زندگی نیاز دارند.28 ].
جدول 1. ویژگی های جمعیت پویا (DPFs) پاسخ جمعیت.
ما سیستم بیماری لایم را به عنوان یک مطالعه موردی بررسی میکنیم و I. scapularis را شبیهسازی میکنیمپویایی جمعیت در شرق ایالات متحده با استفاده از دادههای آب و هوایی مدلسازیشده، و DPFهای کلیدی را برای هر مرحله از زندگی کنهها بهصورت مکانی مشخص، تحلیل و تجسم میکند. ما DPFها را از دینامیک شبیهسازیشده در شرایط آب و هوایی فعلی بررسی میکنیم و آنها را با دادههای مشاهدهشده مقایسه میکنیم تا مشخص کنیم کدام ویژگیها سطوح فعلی خطر بیماری را بهتر پیشبینی میکنند. سپس DPFها را تحت دو سناریو اقلیمی آینده طرحریزی میکنیم و ژئوتصویرسازیهای کلیدی دینامیک بردار پیشبینیشده را در محدوده فضایی ارائه میکنیم. ما با توصیف و تجسم DPFها نشان میدهیم که چگونه میتوانیم تعیین کنیم کدام ویژگیهای جمعیتی خطر بیماری را در شرایط فعلی پیشبینی میکنند و سپس میتوانیم بررسی کنیم که چگونه شرایط آینده ممکن است منجر به تغییر در همین DPFها در آینده شود. ما DPF ها را در زمینه I. scapularis تجزیه و تحلیل می کنیمو خطر بیماری لایم، اما توجه داشته باشید که این رویکرد برای سایر ارگانیسم ها و سیستم های بیماری امیدوار کننده است.
2. روش ها
2.1. روششناسی مدلسازی
تجزیه و تحلیل فراگیر شامل چهار مرحله کلیدی بود. ابتدا، یک مدل جمعیتی قطعی و پویا، بهطور موازی، در یک منطقه جغرافیایی بزرگ اجرا شد تا شبیهسازیهای فضایی صریح تراکم جمعیت در پاسخ به تغییرات دما ایجاد شود. یک گام زمانی روزانه در ارتباط با کوچکترین اندازه سلول شبکه ای که داده های دما از یک مدل گردش جهانی در دسترس بود استفاده شد. دوم، دینامیک جمعیت شبیهسازیشده در هر سلول شبکه برای هر مرحله حیات بردار تحت سناریوهای آب و هوایی فعلی و آینده ثبت شد، و اینها از نظر ویژگیهای جمعیت پویا (DPF) مشخص شدند، که برای برجسته کردن روند جمعیت، فصلی یا ترکیبی انتخاب شدند. از هر دو. سوم، DPF ها برای توانایی آنها در پیش بینی توزیع فعلی ناقل ها یا خطر بیماری های انسانی مورد ارزیابی قرار گرفتند. با استفاده از داده های در دسترس عموم در نهایت، مقادیر DPF که تعیینکنندههای مهم توزیعهای ناقل یا بیماری هستند، در سراسر حوزه فضایی برای طیف وسیعی از سناریوهای آب و هوایی آینده مشاهده شدند. در ادامه هر یک از این مراحل را با کاربرد بیماری لایم به تفصیل شرح می دهیم.
2.2. مدل لایم
یک مدل چرخه زندگی دوازده مرحلهای از کنههای گوزن پا سیاه ( I. scapularis ) (شرح شده در [ 28 )]) برای شبیه سازی با کارایی بالا با استفاده از Simulink (نسخه 7.0) و Matlab (R 2011b) اقتباس شد و در یک خوشه از 48 گره اجرا شد. این مدل با استفاده از میانگین دماهای روزانه (به جای ماهانه) اجرا شد، و مدل به گونهای کدگذاری شد که صریح باشد و بهطور موازی در هر سلول شبکه در یک دامنه فضایی بزرگ اجرا شود. محرکهای دما جمعیت کنههای شبیهسازیشده را از طریق توابع درجه روز، که تاخیرهای توسعه را مدل میکنند، و از طریق پارامترهای فعالیت وابسته به دما، که رفتارهای جستجوی میزبان را مدل میکنند، شکل میدهند (به اطلاعات تکمیلی، جدول S1 مراجعه کنید). با استفاده از یک گام زمانی روزانه، شبیهسازیهای مدل در هر سلول بر روی دامنه تحت هر دو دوره آبوهوای پایه و پیشبینیشده انجام شد. خروجیهای سری زمانی، روزانه ثبت میشوند، شامل جمعیتهای جستجوگر بالغ (QA)، پورههای جستجوگر (QN)، و لارو جستجوگر (QL).اکولوژی Ixodes (به عنوان مثال، [ 34 ، 35 ])، جمعیت ها بین سلول های شبکه تعامل ندارند ( به عنوان مثال ، مهاجرت / مهاجرت مدل سازی نشده است).
2.3. داده های دامنه و آب و هوا
شبیهسازی و تجزیه و تحلیلهای بعدی در شرق ایالات متحده در دامنهای از سلولهای شبکه ۴×۴ کیلومتری انجام شد. داده های آب و هوایی سفارشی در سراسر این شبکه از مدل آب و هوای منطقه ای (تحقیق و پیش بینی آب و هوا (WRF) 3.2.1) شبیه سازی شده در آزمایشگاه ملی دانشگاه تنسی/اوک ریج [ 36 ] به دست آمد. دمای روزانه در هر سلول بهعنوان میانگین دمای حداقل و حداکثر روزانه تولید شده توسط شبیهسازی آب و هوا برای یک دوره زمانی پایه (2001-2004) و دو سناریوی پیشبینیشده با شدت متفاوت (برای 2057-2059) محاسبه شد. سناریوهای پیشبینیشده، مسیر تمرکز نماینده (RCP) 4.5 و RCP 8.5، مربوط به افزایش مداوم نیروی تابشی به 4.5 W/m2 ( سناریوی متوسط) و 8.5 W/m2 است .(سناریوی شدید) به ترتیب در سال 2100 [ 37 ].
2.4. ویژگی های پویا پاسخ جمعیت Ixodes به تغییرات فصلی
ویژگی های جمعیت پویا، انتخاب شده برای برجسته کردن فنولوژی و فصلی جمعیت Ixodes ، همانطور که در جدول 1 برای هر سال توضیح داده شده است، تعیین شد. اینها برای مقایسه دینامیک جمعیت شبیه سازی شده برای هر مرحله زندگی در سطح سلول شبکه برای سه سال شبیه سازی تحت شرایط آب و هوایی پایه و پیش بینی شده مورد استفاده قرار گرفتند. به استثنای میانگین یا میانه ، که به عنوان میانگین یا میانه سه ساله جمعیت روزانه شبیه سازی شده محاسبه می شود، DPF ها برای هر سال از دوره شبیه سازی محاسبه شده و میانگین برای تولید یک مقدار DPF نهایی در هر سلول برای هر سناریوی آب و هوایی محاسبه شد.
DPFهای پاسخ جمعیتی شامل میانگین سه ساله و میانه سه ساله بود. حداکثر جمعیت در طول هر سال ( پیک جمعیت )؛ و میانگین تعداد حداکثرهای محلی در سال ( Peaks per Year ). یک پنجره متحرک 90 روزه برای شناسایی هر قله محلی در طول سال استفاده شد. به منظور شناسایی پیک ها در 90 روز اول خروجی شبیه سازی، 89 روز آخر خروجی شبیه سازی به خروجی اضافه شد تا یک پنجره 90 روزه ارائه شود. به طور مشابه، 89 روز اول خروجی شبیهسازی برای کمک به شناسایی پیکها در 90 روز پایانی شبیهسازی به شبیهسازی اضافه شد.
DPF های فصلی، که برای هر مرحله زندگی تعریف شده بودند، شامل دو کمیت طول فصل و یکی از زمان بندی فصل بودند. یکی از طبقه بندی طول فصل، که IP به IP نامیده می شود ، به عنوان تعداد روزهای بین نقاط عطف در دو طرف حداکثر جمعیت سالانه تعریف شد. نقاط عطف به عنوان تغییرات در تقعر سری زمانی جمعیت صاف شده با لس (پنجره 30 روزه) تعریف شد. تغییرات در تقعر با استفاده از یک معادله اختلاف مرکزی 3 نقطه ای در سری زمانی هموار تعیین شد. کمیت طول فصل دوم به عنوان تعداد روزها از حداکثر جمعیت سالانه تا حداقل جمعیت سالانه تعریف شد که به آن Peak to Trough گفته می شود. زاویه موجممکن است به عنوان زمان نسبی فصل هر سلول درک شود. برای تعیین زاویه موج ، تحلیل موجک پیوسته با استفاده از موجک مورلت [ 38 ] برای دوره حداکثر توان، 90.5 روز انجام شد (به اطلاعات تکمیلی مراجعه کنید). DPFهای ترکیب فصلی و جمعیت مطلق شامل IP Pop تعداد روزهای تیک در طول فصل هر مرحله زندگی است (یعنی جمع جمعیت کنه برای تمام روزهایی که در محاسبه IP به IP گنجانده شده است). علاوه بر این، UQ/IQR برای تخمین دوره در سال تقویمی که در آن بیشترین جمعیت شبیه سازی شده چارک رخ می دهد، استفاده می شود. بنابراین، UQ/IQR با انتخاب نقاط زمانی (روزها) که در آن جمعیت چارک بالایی رخ می دهد، سپس میانگین محدوده بین چارکی این نقاط زمانی را انتخاب می کنیم ( جدول 1 ).
2.5. مقایسه DPF ها با داده های مشاهده شده
DPFهای به دست آمده از مدل همانطور که در بالا توضیح داده شد برای حضور I. scapularis در سطح شهرستان مشاهده شده (کدگذاری شده در سه سطح به عنوان غایب، گزارش شده و ایجاد شده) و بروز بیماری لایم (که در چهار سطح به عنوان هیچ / حداقل، کم، متوسط و زیاد کدگذاری شده است) مناسب بودند. ) از مراکز کنترل و پیشگیری از بیماری (CDC) به دست آمده است [ 33 ، 39 ]. برای همه تحلیلها، چهار طبقهبندی گزارششده در همه تقسیمبندیهای ممکن گروهبندی شدند (به عنوان مثال، برای بیماری لایم، دوگانگیسازیها شامل حداقل/هیچ در مقابل کم، متوسط و زیاد؛ حداقل/هیچ و کم در مقابل متوسط و زیاد؛ و حداقل/هیچ، کم و متوسط در مقابلبالا). DPFها از نظر مکانی به سطح شهرستان میانگین داده شدند و با داده های مشاهده شده (CDC) با استفاده از هر دو ناحیه زیر منحنی مشخصه عملکرد گیرنده (AUC) و رگرسیون لجستیک برای تعیین توانایی پیش بینی هر DPF مقایسه شدند. AUC (محدوده: 0 تا 1) دقت پیشبینی مدل را برای یک نتیجه دوگانه کوانتیزه میکند، جایی که مقدار 0.5 نشاندهنده عدم توانایی پیشبینی، مقدار 1 نشاندهنده تمایز کامل و مقدار 0 نشاندهنده عدم تمایز است. برای ارزیابی تغییرات فضایی بالقوه در توانایی DPFs برای پیشبینی خطر بیماری لایم، AUC برای DPFهای منتخب نیز برای شهرستانها در سه منطقه (شمال شرقی، جنوب و غرب میانه)، همانطور که توسط سرشماری ایالات متحده تعریف شد، تعیین شد.
2.6. تجزیه و تحلیل حساسیت فضایی
برای ارزیابی حساسیت DPF به خود همبستگی فضایی، خروجیهای مدل تعریفشده در بالا نیز با دادههای شیوع بیماری لایم و کنه با استفاده از هر دو مدل لجستیک (غیر فضایی) و مدل لجستیک فضایی مناسب بودند. مدل رگرسیون فضایی به صورت زیر تعریف می شود:
که در آن y i رده بیماری لایم مشاهده شده تقسیم شده است، α خطر کلی پایه است، λi اثرات تصادفی فضایی خاص شهرستان است و β نشان دهنده نسبت شانس ورود به سیستم مرتبط با DPFهای پاسخ جمعیت است ( Xi ) . این مدل از کنترلهایی برای اثرات خودهمبستگی فضایی با استفاده از مدل خودرگرسیون شرطی ذاتی (CAR) استفاده کرد [ 40 ]. مدل CAR، که اغلب با توزیع شرطی λi ، با توجه به همسایگانش فرموله می شود، فرض می کند که λi ، برای هر شهرستان، i، میانگین فضایی همسایگان آن است. توزیع شرطی گاوسی است، با میانگین و واریانس τ2 ∕ m i ، که در آن i~i نشاندهنده این است که شهرستان، i ، مرز مشترکی با شهرستان i » دارد، و m i تعداد همسایگان اشتراکگذاری مرز برای شهرستان i است. درصد تغییر در β و مقدار پارامتر، τ2 ، که درجه فضایی را به طور مشابه کنترل میکند، برای ارزیابی بهبود تناسب ارائه شده با افزودن عبارت فضایی به مدل رگرسیون لجستیک استفاده شد.

3. نتایج
دینامیک جمعیت شبیهسازیشده برای سه مرحله حیات کنههای جستجوگر از یک مدل دما اجباری شبیهسازیشده در شبکهای از سلولهای ۴×۴ کیلومتری که شرق ایالات متحده را تحت هر دو آب و هوای پایه و پیشبینیشده پوشش میدهد، به دست آمد. بر اساس این پویاییهای جمعیت شبیهسازیشده، ده ویژگی جمعیت پویا (DPF) مشتق شده و بهصورت مکانی تجزیه و تحلیل شدند تا پاسخ جمعیت به اقلیم فعلی و آینده در سراسر حوزه را مشخص کنند. همبستگی های دوتایی بین DPF ها تعیین شد و هر DPF تحت شرایط آب و هوایی فعلی برای توانایی آن در تشخیص خطر بیماری لایم و حضور/غیاب ناقل با استفاده از داده های مشاهده شده از CDC ایالات متحده ارزیابی شد.
3.1. همبستگی بین DPF ها
برای تعیین درجه خطی بودن DPFها و بنابراین، برای تعیین کمیت جنبههای مرتبط منحنی جمعیت، همبستگیها برای همه جفتهای DPF محاسبه شد. همبستگی قوی بین DPFهای مختلف جمعیت مطلق ( میانگین ، میانه ، و جمعیت اوج ) در تمام مراحل زندگی جستجوگر بر روی وسعت فضایی تجزیه و تحلیل وجود داشت ( rs ≥ 0.96؛ جدول 2 را ببینید ). پیک ها در سال و IP Pop نیز با DPFهای جمعیت مطلق و یکدیگر در مرحله جستجوی بزرگسالان همبستگی زیادی داشتند (r s ≥ 0.88) در حالی که پیک در سال نیز همبستگی بالایی داشت (rs ≥ 0.90)، با جمعیت مطلق DPF و IP به IP در مراحل پوره و لاروی جستجو. فقط زاویه موج با تمام DPFهای دیگر در مراحل پوره و بالغ همبستگی معکوس داشت. به استثنای همبستگی آن با UQ/IQR (QA: rs = -0.77 ؛ QN: rs = -0.81 ) ، این رابطه معکوس ضعیف بود ( rs ≥ -0.63) . در مرحله لاروی جستجو، فقط IP به IP یک همبستگی معکوس ضعیف با سایر DPFها نشان داد (به جز همبستگی آن با UQ/IQR ). هیچ DPF زمانبندی قویاً با هیچ DPF زمانبندی دیگری در هیچ مرحلهای از زندگی مرتبط نبود.
جدول 2. ضرایب همبستگی اسپیرمن (r s ) * بین DPFها در هر سلول برای هر مرحله حیات جستجو تحت سناریوی آب و هوای پایه ارزیابی شد.
3.2. مقایسه DPF ها با داده های مشاهده شده
هر DPF برای حضور کنه مشاهده شده یا خطر بیماری لایم از CDC ایالات متحده مناسب بود و یک مقدار AUC تولید میکرد، یک شاخص تبعیضآمیز که امکان مقایسه پیشبینیهای مداوم با مشاهدات دوگانه را بدون نیاز به نقاط برش ذهنی فراهم میکرد. ماه اوج و جمعیت اوج بیشترین توانایی تبعیض آمیز را در تمام مراحل زندگی در مقایسه با داده های خطر بیماری CDC Lyme نشان دادند ( جدول 3 ؛ AUC = 0.54 تا 0.90). در میان اکثر DPFها، دوقطبیسازیهای حداقل در مقابل زیاد یا حداقل/کم/متوسط در مقابل بالا مقادیر AUC بالاتری نسبت به تقسیمبندیهای باقیمانده دادند. دوگانگی حداقل در مقابل.پایین/متوسط/بالا بدترین عملکرد را در آنالیزهای AUC در تمام مراحل زندگی جستجوی انجام داد. Peak to Trough ، IP به IP ، IP Pop ، UQ/IQR ، و Wave Angle توانایی پیشبینی متناقضی را در هر سه مرحله زندگی نشان دادند. از این میان، فقط IP Pop در مرحله حیات QN توانایی تشخیصی بهبود یافته را نشان داد.
مقایسه DPFs با داده های حضور کنه CDC توانایی تبعیض کمتری را نشان داد. جمعیت پیک، ماه اوج، اوج تا پایین و UQ/IQR پیشبینیکنندههای آماری معنیداری برای حضور کنه در تمام مراحل زندگی جستجو و تقسیمبندی دادههای CDC بودند، اما مقادیر AUC پایین بود و از 0.53 تا 0.71 برای QA، از 0.54 تا 0.69 برای QA متغیر بود. QN و از 0.52 تا 0.69 برای QL. برای زاویه موج QL و حضور تیک، AUC از 0.66 تا 0.7 متغیر بود. همراه با ماه اوج QA و جمعیت اوج QA ، این یکی از قویترین پیشبینیکنندههای حضور کنه بود. با این حال، برای دوگانگی حداقل/هیچ در مقابل کم/متوسط/بالا، DPFs، میانگین ،میانه ، پیک در سال ، IP به IP ، IP Pop و UQ/IQR ، به طور یکنواخت در تمام مراحل زندگی جستجوی معنی دار نبودند. مقادیر AUC ماه اوج برای QN و QL بسیار مشابه و برای QA کمی کمتر بود.
جدول 3. تجزیه و تحلیل ناحیه زیر منحنی مشخصه عملکرد گیرنده (AUC) که ویژگی های جمعیت پویا (DPFs) را با بروز بیماری لایم مشاهده شده و حضور کنه مقایسه می کند.
3.2.1. تحلیل های منطقه ای
یک تجزیه و تحلیل منطقه ای، مشابه با تجزیه و تحلیل AUC قبلی، برای ماه اوج و جمعیت پیک انجام شد تا میزان توانایی تبعیض آمیز را بر اساس مکان مشخص کند. در بیشتر موارد، AUC برای ماه اوج و جمعیت پیک بالاترین ( جدول 4 ) در منطقه غرب میانه شرق ایالات متحده (AUC: 0.80 تا 0.96)، جایی که AUC اغلب از نظر آماری معنی دار بود، مشاهده شد. مقادیر AUC در شمال به طور مداوم کمتر از غرب میانه (AUC: 0.71 تا 0.78)، برای هر دو DPF بود. در جنوب، ماه اوج معمولاً توانایی پیشبینی آماری معنیداری بالاتری را برای QN و QL نسبت به جمعیت اوج نشان میدهد.در حالی که هر دو DPF توانایی پیشبینی مشابهی را برای QA نشان دادند.
جدول 4. تحلیل های فرعی AUC * منطقه ای.
3.2.2. تجزیه و تحلیل حساسیت فضایی
ضرایب رگرسیون تولید شده در مدلهای خودرگرسیون شرطی برای هر DPF در هر مرحله زندگی در دادههای CDC تفاوت اساسی با ضرایب تولید شده توسط یک رگرسیون غیر فضایی (نشان داده نشده) نداشت. مدلهای خودرگرسیون شرطی مقادیر کم τ2 ( 0.003، 0.06) را نسبت به رهگیری و/یا مقدار پارامتر نشان دادند، که نشان میدهد سهم خودهمبستگی فضایی کوچک است.
3.3. تغییرات در توزیع جغرافیایی DPFs
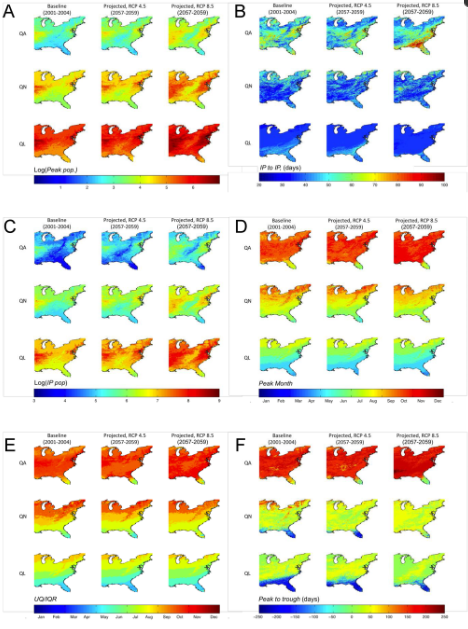
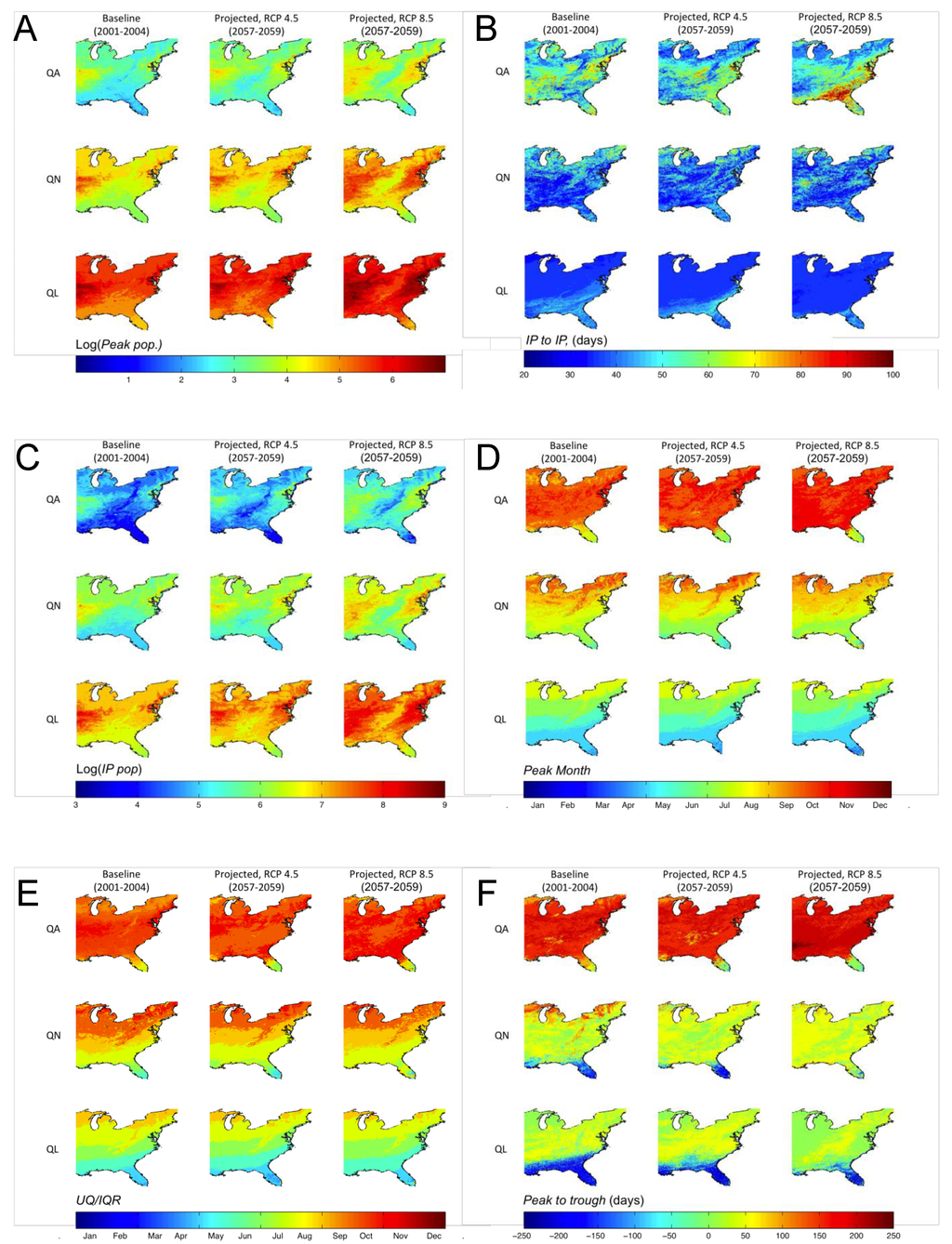
DPFها در تمام سلولها (N = 262152) تحت سناریوهای آب و هوایی آینده به طور قابل توجهی ( P <0.001) متفاوت از شبیهسازیهای پایه بودند. میانگین، میانه و اوج جمعیت شبیهسازیشده، همگی افزایشهایی را در بیشتر مناطق شرقی ایالات متحده نشان میدهند، با بیشترین افزایش در سناریوی RCP 8.5. به طور خاص، 18.8٪، 7.7٪ و 4.1٪ از سلول ها به ترتیب در سناریوی RCP 8.5 افزایش یک مرتبه یا بیشتر در اوج جمعیت QA، QN و QL نشان دادند. نسبت به خط پایه، مناطق با بالاترین میانگین پیش بینی شده، میانه و جمعیت کنه اوج هم به سمت شمال و هم به سمت جنوب گسترش یافتند تا به مناطقی با مقادیر کم DPF که در سراسر رشته کوه آپالاچی رخ می دهد تجاوز کنند (به عنوان مثال، شکل 1 ).
در حالی که هر دو سناریوی پیشبینیشده میانگین پیکهای مرحله زندگی جستجوی شبیهسازی شده در سال را نشان دادند که به طور قابلتوجهی با حالت پایه متفاوت بود و دو سناریوی پیشبینیشده به طور قابلتوجهی با یکدیگر متفاوت بودند، هیچ تغییر اساسی (01/0 پیک در سال) در زمانی که این مقایسهها انجام شد، وجود نداشت. در کل دامنه تغییرات در پاسخ جمعیت بین سناریوها، همانطور که توسط پیک در سال نشان داده شده است، عدم پاسخگویی یکنواخت را نشان می دهد ( شکل 1). تعداد سلولهایی که پیشبینی میشود دو یا چند پیک جمعیت در سال برای مرحله زندگی QA داشته باشند در مناطق شمالی افزایش یافت، در حالی که کاهش خالص در تعداد سلولهای با بیش از یک پیک در مناطق جنوبی وجود داشت. در مقابل، برای مراحل زندگی QL و QN، تعداد سلولهایی که بیش از یک پیک جمعیت را در سال تجربه میکنند به طور یکنواخت در سراسر حوزه فضایی کاهش یافت.
در سناریوهای RCP 4.5 و RCP 8.5 در مقایسه با خط پایه، طول فصل برای QA، همانطور که توسط IP به IP تعریف شده است، افزایش قابل توجهی از فصول 50-70 روزه به 60-90 و 80-100 روز در جنوب دارد. . در شمال، یک کاهش خالص در طول فصل QA برای هر دو سناریو آینده وجود دارد ( شکل 1). برای QN، طول فصل پیشبینیشده همان خط پایه باقی ماند، اگرچه کاهش جزئی در طول فصل برای بیشتر بخش شمالی منطقه مورد مطالعه در هر دو سناریو پیشبینیشده، و افزایش تقریباً 40 روزه در طول فصل در بخش کوچکی وجود داشت. از غرب میانه محصور در خشکی. طول فصل QL پیش بینی شده کاهش تقریباً 10 روزه را در مناطق جنوبی نشان داد، در حالی که منطقه جغرافیایی کلی با طول فصل بالاتر با افزایش شدت سناریو کاهش یافت. تغییرات در طول فصل پیش بینی شده، همانطور که توسط Peak to Trough تعریف شده است ، تقریباً در سراسر دامنه یکنواخت بود (نشان داده نشده است). QA Peak to Trough به طور متوسط هشت روز در RCP 4.5 و 1.4 روز در RCP 8.5 در مقایسه با پایه افزایش یافت. QN و QL Peak to Troughدر سناریوی RCP 8.5 به ترتیب 56.6 و 31.1 روز و در سناریوی RCP 4.5 به ترتیب 15 و 10 روز کوتاه شد.
شکل 1. ( الف ) گزارش جمعیت پیک ، ( B ) IP به IP ، ( C ) گزارش IP Pop ، ( D ) ماه اوج ، ( E ) UQ/IQR و ( F ) اوج به پایین برای جستجوی بزرگسالان (QA) ، پوره های جستجوگر (QN) و لاروهای جستجوگر (QL).
نوردهی DPF، IP Pop ، که تعداد روزهای تیک را در طول یک فصل با نقاط عطف حول حداکثر جمعیت سالانه شمارش میکند، شباهت کمی به نقشههای IP به IP دارد ( شکل 1 ). با افزایش شدت سناریوهای پیش بینی شده، IP Pop در زمان قرار گرفتن در سراسر دامنه افزایش می یابد. مناطقی که بیشترین تعداد کنههای پایه را دارند، که در طول فصل تعیینشده نقطه عطف وجود دارند، در هر دو جهت شمال و جنوب و مرکز اطراف غرب میانه و شمال شرق به سمت خارج گسترش مییابند.
نتایج زاویه موج (نگاه کنید به اطلاعات تکمیلی؛ شکل S1) نشان داد که پویایی تمام مراحل زندگی تحت هر دو شرایط آب و هوایی پیشبینیشده، ≤4 روز در سراسر حوزه شبیهسازی شده از شرایط آب و هوایی پایه عقب است. ماه پیشبینیشده جمعیت اوج ( ماه اوج ) و UQ/IQR برای QA عموماً به ماههای قبلی منتقل شد. با این حال، با افزایش شدت سناریو، QN و QL به طور کلی به ماههای بعدی در سراسر منطقه جغرافیایی منتقل شدند.
نتایج زاویه موج (نگاه کنید به شکل S1) نشان داد که دینامیک تمام مراحل زندگی تحت هر دو شرایط آب و هوایی پیشبینیشده، 4 روز در سراسر حوزه شبیهسازی شده از آب و هوای پایه عقبتر است. ماه پیشبینیشده جمعیت اوج ( ماه اوج ) و UQ/IQR برای QA عموماً به ماههای قبلی منتقل شد. با این حال، با افزایش شدت سناریو، QN و QL به طور کلی به ماههای بعدی در سراسر منطقه جغرافیایی منتقل شدند.
4. بحث و نتیجه گیری
هنگام بررسی واکنش جمعیت های ناقل به تغییرات آب و هوایی، تغییرات در فنولوژی، فصلی و سایر ویژگی های دینامیکی را می توان در محدوده فضایی و مراحل زندگی ارگانیسم مورد نظر پیش بینی کرد. خطر VBD هم به زمان و هم به احتمال قرار گرفتن در معرض بردار بستگی دارد، و بنابراین، مشخص کردن پاسخ پویای جمعیت در فضا برای پیشبینی و مدیریت ریسکهای بالقوه آینده بسیار مهم است. در اینجا، با استفاده از شبیهسازی صریح فضایی یک مدل جمعیت مبتنی بر اقلیم و ساختار مرحلهای، چارچوبی را برای ارزیابی اثرات استاتیک و پویا تغییرات آب و هوا بر روی جمعیتها در مناطق جغرافیایی بزرگ ارائه میکنیم.
یافته های ما با توجه به Ixodes scapularis هم روش و هم کاربرد آن را نشان می دهد. استخراج و تحلیل ویژگیهای جمعیت پویا کلید رویکرد تحلیلی است. DPF ها اطلاعات کمی را در مورد طیف وسیعی از ویژگی های جمعیت ارائه می دهند و امکان مقایسه بین خروجی شبیه سازی پویا و داده های بیماری مشاهده شده و همچنین بین آب و هوای پایه و پیش بینی شده را فراهم می کنند. DPFهای مطلق، مانند میانگین ، میانه و اوج جمعیت را می توان به عنوان شاخص های بقا تفسیر کرد، در حالی که DPF های زمان بندی، مانند تعداد روزهای از حداکثر جمعیت سالانه تا حداقل جمعیت سالانه ( پیک تا پایین ) و ماهی که در آن اوج جمعیت رخ می دهد. (ماه اوج ) زمان و طول فصل یک مرحله زندگی معین را مشخص می کند.
در مورد Ixodes ، DPF های مرتبط با اوج منحنی جمعیت شبیه سازی شده، پیک جمعیت و ماه اوج ، مهم ترین در پیش بینی خطر بالای بیماری لایم هستند، اگرچه همه DPF ها سطحی از توانایی تبعیض را نشان دادند. تجزیه و تحلیل AUC نشان داد که دوگانگی جداسازی ریسک بالا توانایی تمایز را در تمام DPF ها و مراحل زندگی بهبود می بخشد. تجمیع ریسک متوسط و بالا همچنین توانایی تشخیصی بهبود یافته در مراحل زندگی و DPFها را در مقایسه با حداقل در مقابل نشان داد.دوقطبی شدن کم/متوسط/بالا. این روند بهبود، از آنجایی که خطر بیماری بالا به تدریج در یک دسته واحد جدا می شود، به شدت نشان می دهد که این DPF ها در پیش بینی زمان و مکان خطرات بالاتر بیماری لایم مفید هستند.
هنگامی که DPF ها برای دو سناریو آب و هوایی پیش بینی شده بررسی می شوند، نشان می دهیم که پاسخ جمعیت پویا I. scapularis در طول مراحل زندگی یکنواخت نیست و در فضا متفاوت است. تغییرات مکانی در ویژگیهای زمانی شامل جابجاییهای جغرافیایی در فصل میشود، و این جابهجاییها آنطور که به طور شهودی فرضیه میشود، پیوسته به سمت شمال نیستند. در حالی که ماهی که در آن بیشترین تعداد کنه ها در حال جستجو هستند ( ماه اوج ) برای مرحله زندگی بزرگسالی به تعویق افتاده است ( شکل 1 )، پیک های QN و QL تغییرات جغرافیایی یکنواخت به فصل جستجوی اولیه را نشان نمی دهند. همچنین، Peak to Trough و IP to IP نشان دهنده تغییرات احتمالی در طول فصل در سناریوهای پیش بینی شده است. تغییرات فضایی در DPFهای مطلق، ماننداوج جمعیت ، بر اساس منطقه متفاوت است. به عنوان مثال، انتظار میرود که جمعیت قلهها در مناطق غرب میانه و شمال شرق هر دو در مقایسه با رشته کوه آپالاچی یا ساحل خلیج، جایی که انتظار میرود این جمعیتها پایدارتر باقی بمانند، بسیار بیشتر افزایش یابد.
اگرچه یافتهها مبنی بر اینکه ماه اوج QL و جمعیت اوج توانایی پیشبینی بالایی برای خطر بیماری لایم نشان میدهند، قابل توجه است، پیامدهای علی این یافته و سایر موارد مشابه آن، باید با احتیاط تفسیر شوند. خطر بیماری مستقیماً با مرحله جستجوی لارو مرتبط نیست، که اولین وعده غذایی خون را در چرخه زندگی می گیرد، و بنابراین، تنها در شرایط نادری که لاروها از طریق تخمدان آلوده می شوند، مسئول انتقال لایم است. به همین ترتیب، ماه اوج QN و اوج جمعیتدارای مقادیر AUC مشابهی برای همه تقسیمبندیهای خطر بیماری لایم هستند، اثری که عمدتاً ناشی از شباهت پاسخ کنه به دما در این دو مرحله زندگی است، نه روابط علّی متقابل با بیماری. روابط زمانی پیچیده در این جمعیت ها ذاتی است: به عنوان مثال، پوره های جستجوگر و لاروهای جستجوگر تقریباً در همان زمان از سال به اوج خود می رسند، و جمعیت آنها در یک مکان مشخص ظاهراً همبستگی دارند، اگرچه جمعیت QN از جمعیت QL در همان سال، اما QL سال های قبل.
همانند سایر تحلیلهای مدلسازی بومشناختی، کیفیت دادهها سودمندی این چارچوب تحلیل را برای یک سیستم معین تعیین میکند. در تجزیه و تحلیل ما، کیفیت داده های CDC ممکن است برای فقدان AUC قابل توجه DPF ها در مقایسه با داده های کنه مشاهده شده باشد. دادههای حضور/غیاب کنه با استفاده از روشهای مختلف، مانند کشیدن و بررسی گوزنها، اغلب تحت محدودیتهای جدی منابع جمعآوری میشوند [ 33 ]]. به جای ارائه اطلاعات منسجم و سیستماتیک در مورد حضور و عدم حضور کنه، مجموعه داده ملی کنه یک دسته بندی درشت برگرفته از اطلاعات متفاوت ارائه می دهد. این برخلاف مجموعه داده ملی بیماری لایم است که بر اساس یک استاندارد گزارش دهی سازگار است. با توجه به کیفیت بالاتر داده های جمع آوری شده، این مجموعه داده در اثبات نتایج مدل ما مفیدتر است.
سایر عوامل آب و هوایی به غیر از دما، مانند رطوبت، بر روی Ixodes spp تأثیر می گذارند . فعالیت [ 41 ، 42 ] و با خطر بیماری لایم انسانی [ 43 ] ارتباط دارد. مدل جمعیتی مورد استفاده در اینجا پاسخ Ixodes به رطوبت را در بر نمیگیرد، و اگرچه دادههای جمعیت شبیهسازیشده ما مطابقت خوبی با بروز Lyme نشان میدهد، این امکان وجود دارد که شامل سایر متغیرهای کلیدی محیطی ممکن است مطابقت بیشتری داشته باشد. به همین ترتیب، جمعیت میزبان و پاتوژن در تجزیه و تحلیل ما، که به پویایی ناقل محدود بود، در نظر گرفته نشد. تحقیقات نسبتا کمی در مورد پاسخ های بالقوه جمعیت Borrelia انجام شده استspp _ تحت شرایط آب و هوایی تغییر یافته با این حال، پیشنهاد شده است که تغییرات در فنولوژی Ixodes در پاسخ به تغییرات آب و هوایی ممکن است بر تکامل پاتوژن های مختلف منتقله از کنه تأثیر بگذارد، به طوری که طول عمر، انتقال و بیماری زایی آنها را اصلاح کند [ 44 ]. دینامیک میزبان همچنین می تواند بر تراکم ناقل آلوده و در نتیجه خطر انسانی در انواع سیستم های VBD تأثیر زیادی بگذارد [ 45 ، 46 ، 47 ]. در مورد بیماری لایم، فراوانی میزبانهای کلیدی، مانند موش و سنجاب، نشان داده شده است که تراکم پورههای آلوده را در جنگلهای برگریز شرقی پیشبینی میکند [ 48 ].]؛ در مناطق دیگر، مانند جنوب ایالات متحده، اعتقاد بر این است که مارمولک ها به دلیل صلاحیت ضعیف میزبان یا اثرات پیشگیرانه حیوانی، اثر کاهش دهنده ای بر گسترش بیماری لایم دارند [ 49 ، 50 ]. گنجاندن دینامیک میزبان، ناقل و پاتوژن در یک مدل ترکیبی، چالشهای روششناختی و محاسباتی قابلتوجهی ایجاد میکند، اما همچنین احتمالاً به درک مکانیکی ما از تغییر ریسک VBD در شرایط محیطی آینده کمک زیادی میکند. ما توجه می کنیم که یک شبیه سازی، خلاصه سازی (به عنوان مثال، DPFs) و رویکرد تجزیه و تحلیل مشابه را می توان با چنین مدل ترکیبی دنبال کرد. با این حال، سایر خلاصهسازیها (به عنوان مثال، R0 ) برای تصویرسازی جغرافیایی در آن زمینه در دسترس هستند (به عنوان مثال، [ 51 , 52 )]).
مشارکت روششناختی که توسط تحلیل مدلسازی شرح داده شده در اینجا قابل توجه است. ما یک ارزیابی کمی از پویایی جمعیت – با عواقب بالقوه برای خطر بیماری – در شرایط آب و هوایی آینده ارائه میکنیم که با استفاده از یک مدل مکانیکی صریح فضایی ممکن میشود [ 53 ]. توصیف فضایی ما از DPF ها امکان ارزیابی بصری دقیق را فراهم می کند (به عنوان مثال، شکل 1در کنار تجزیه و تحلیل کمی، پاسخ پویای جمعیت به آب و هوای آینده، تغییرات بالقوه ای را آشکار می کند که غیر شهودی هستند. به عنوان مثال، در سراسر شرق ایالات متحده، تحت دماهای پیش بینی شده در مقایسه با سناریوی پایه، پوره ها و لاروها در اوایل فصل به اوج جمعیت خود می رسند، در حالی که پیش بینی می شود بزرگسالان در اواخر فصل به اوج جمعیت برسند ( شکل 1). ). رویکرد اتخاذ شده در اینجا همچنین ارزش مدلسازی فراوانی را برجسته میکند، که بر خلاف تناسب زیستگاه یا سایر اقدامات ثابت، امکان بررسی فنولوژی و فصلی بودن در بین مراحل زندگی و پیامدهای بالقوه برای (و همبستگی با) خطر بیماری را فراهم میکند. به عنوان مثال، IP به IPنشان می دهد که طول “فصل” لارو در سه سناریو دما پایدار است، در حالی که مراحل بالغ و تا حدی پوره “فصل” را نشان می دهند که به شدت به دماهای افزایش پیش بینی شده حساس هستند. با استفاده از روشهای سنتی که به عنوان مثال، اثرات سالانه کل را بررسی میکنند، چنین پاسخهای خاص مرحله حیات در زمان و مکان، آشکار نیستند.
ما در بالا نسبت به تفسیر علی قدرت پیشبینی DPF هشدار میدهیم. یک همبستگی قوی بین DPF و بروز بیماری مشاهده شده ممکن است نشان دهنده یک رابطه علّی نباشد، اما چنین یافته ای می تواند فرضیه هایی را مطرح کند که در نهایت منجر به درک مکانیکی بیشتر از رابطه بین جمعیت های ناقل و خطر بیماری در فضا و زمان و بنابراین، بهبود یافته است. درک علت و معلولی در نهایت، مدلهای جمعیتی، مانند مدلی که در اینجا مورد بررسی قرار گرفت، همچنین میتواند برای ارزیابی اثربخشی و صرفهجویی در مداخلات بالقوه سلامت عمومی [ 53 ]، مانند کنترل ناقل یا میزبان (به عنوان مثال، [ 54 ، 55 ، 56 ) استفاده شود.] برای بیماری لایم). تجزیه و تحلیل توأم از اثر دما در حضور یک برنامه کنترل ناقل، گسترش آشکار این رویکرد خواهد بود، و چنین کاربرد این مدل برای بسیاری از ناقلین، مداخلات و بیماری های مختلف امکان پذیر است.
ما توانایی یک مدل جمعیت پویا صریح فضایی را برای تمایز بین ویژگیهای جمعیت پویا که به شدت با خطر بیماری مرتبط است، و همچنین برای توصیف پاسخ جغرافیایی متنوع مراحل زندگی I. scapularis به پویایی آب و هوا نشان دادهایم. استفاده از چنین رویکردی برای توصیف تغییرات در پویایی محدود به بیماری لایم نیست. این تکنیک ممکن است بینش جدیدی در مورد پاسخ های پویا طیفی از ناقلان بیماری به تغییرات محیطی، به ویژه تغییرات در ویژگی های فصلی و فنولوژیکی آنها ارائه دهد. چنین تحلیلهایی ممکن است اطلاعات مفیدی در مورد خطر متعاقب بیماری منتقله از طریق ناقل در شرایط آینده ارائه دهد.



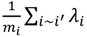
بدون نظر